

Influenza 2019/20 season, Japan
(IASR Vol. 41 p191-193: November 2020)
The 2019/20 influenza season (from week 36 in September 2019 to week 35 in August 2020) was characterized by the predominance of the influenza A/H1pdm09 subtype, and influenza B, mainly Victoria lineage, also increased from week 51 in 2019.
Epidemiology of the 2019/20 influenza season: Under the National Epidemiological Surveillance of Infectious Diseases (NESID) system, approximately 5,000 influenza sentinel sites (approximately 3,000 pediatric and 2,000 internal medicine facilities) report patients diagnosed as influenza on a weekly basis. The number of patients reported per sentinel exceeded 1.0 (the usual indicator of the nationwide start of influenza season) in week 37 (September 9-15) of 2019 due to a local epidemic of mainly influenza A in Okinawa Prefecture (see p.194 of this issue), but the nationwide influenza season was considered to have started in week 45 (November 4-10) of 2019 (Fig. 1 & https://www.niid.go.jp/niid/ja/10/weeklygraph.html).
{kind=link}
In week 52 (December 23-29), the number exceeded the alert level of 10.0 in all 47 prefectures and the peak was also in the same week (23.24 patients/sentinel, which was approximately half the value of the average year’s peak).
Based on the reported sentinel surveillance data, the estimated number of influenza patients who visited medical facilities (between week 36 of 2019 and week 17 of 2020) was approximately 7,289,000, approximately 61% of that during the same period in the previous season (12,005,000). According to hospitalized influenza patient surveillance (data from approximately 500 designated sentinel hospitals with ≥300 beds), the number of hospitalized influenza patients was 12,982, approximately 64% of that during the same period in the previous season. Among patients diagnosed with acute encephalitis (including encephalopathy), a category V notifiable infectious disease, 254 were diagnosed with influenza encephalitis, the largest number in the past three seasons (https://www.niid.go.jp/niid/images/idsc/disease/influ/fludoco1920.pdf).
Nationwide excess deaths in the 2019/2020 season and January-June 2020 when the COVID-19 pandemic started were analyzed by two algorithms used in the US and Europe, estimating 191-4,577 and 319-7,467, respectively (see p.207 of this issue).
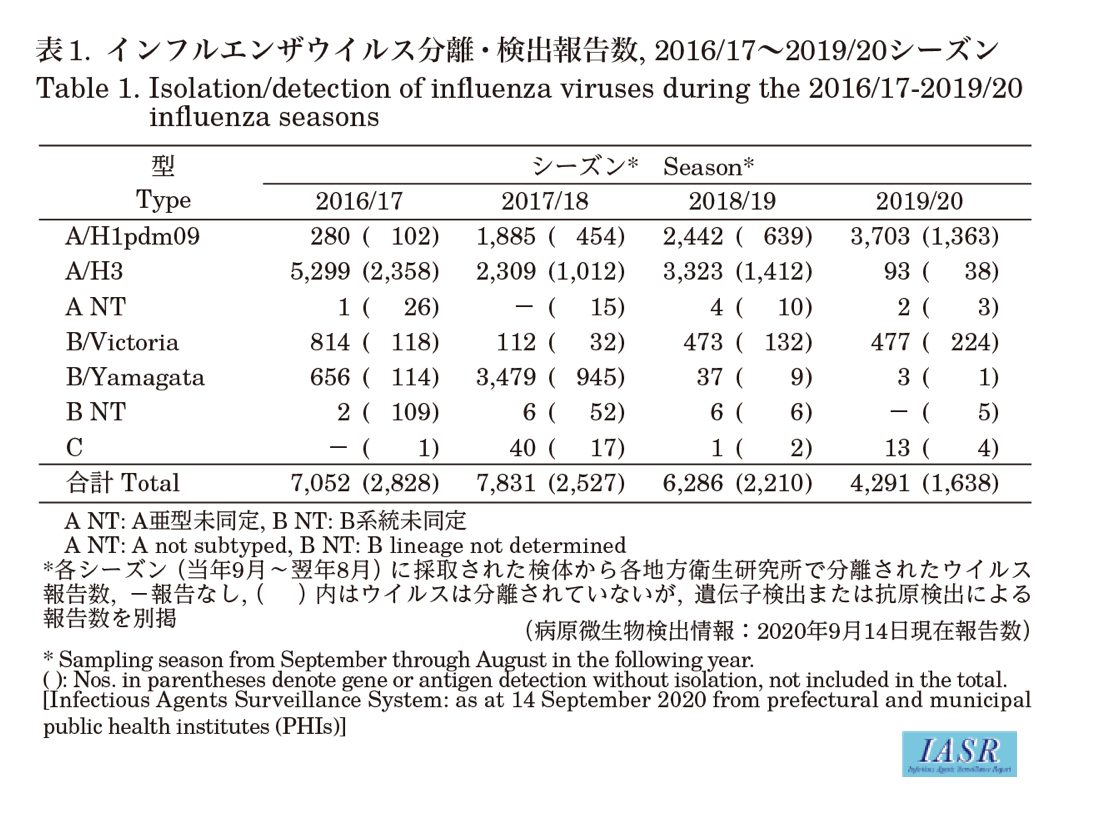
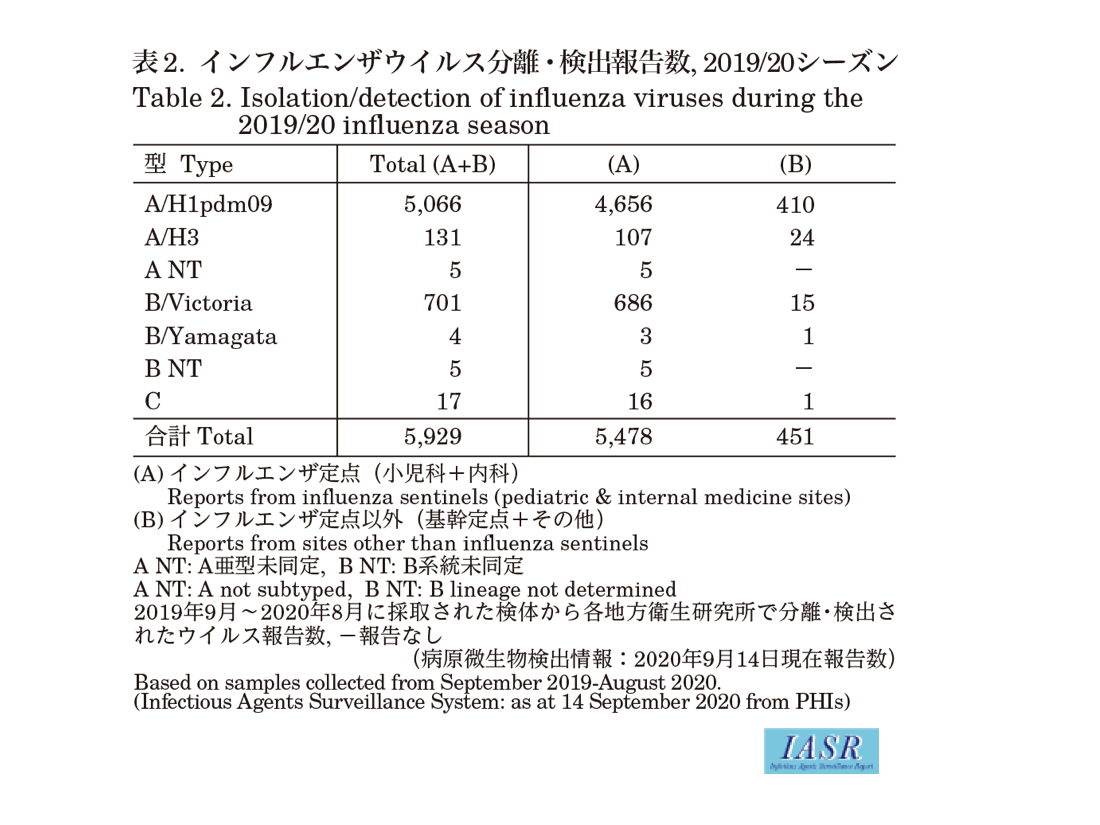
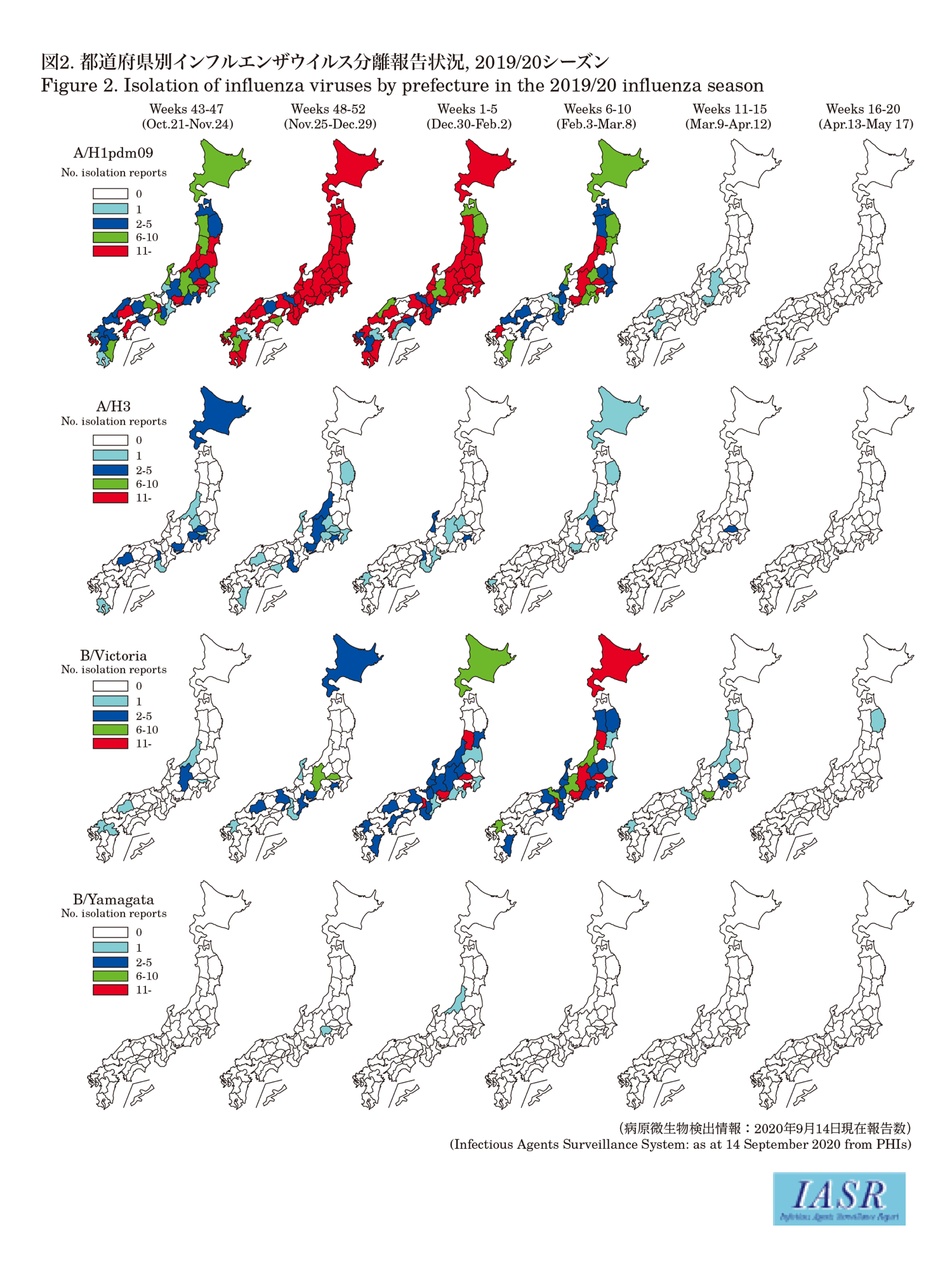
Isolation/detection of influenza virus in the 2019/20 season: Prefectural and municipal public health institutes (PHIs) reported the isolation/detection of 5,929 influenza viruses (4,291 isolates and 1,638 detections without isolation) (Table 1). Among them, 5,478 were reported from influenza sentinel sites and 451 from non-sentinel sites (Table 2). Influenza type A/H1pdm09 comprised 85%, A/H3 comprised 2%, and type B comprised 12% (Victoria lineage 98.7%, Yamagata lineage 0.6%, and unknown lineage 0.7%) (Table 2). Although isolations of A/H1pdm09 increased from week 46 (November 11-17) of 2019, those of type B Victoria lineage started to increase from week 51 (December 16-22) of 2019, surpassing those of type A from week 9 (February 24-March 1) of 2020 (Fig. 1 & Fig. 2). As for the winter season in the temperate region of the southern hemisphere, no epidemic was observed even in May-August when the epidemic usually peaks (see p.208 of this issue).
{kind=link}
{kind=link}
{kind=link}
Genetic and antigenic characteristics of 2019/20 isolates: The National Institute of Infectious Diseases (NIID) conducted genetic and antigenic analyses of isolates from Japan and other Asian countries; antigenic analyses were done using ferret antiserum (p.195 of this issue). The hemagglutinin (HA) genes of all A/H1pdm09 isolates analyzed belonged to genetic clade 6B.1A, and the majority of viruses possessed 183P substitution in HA.
Although A/H3 strains are largely divided into two groups, 3C.2a and 3C.3a, in the HA gene tree, the dominant strains analyzed in the 2019/20 season belonged to 3C.2a, as in the past five seasons. Within genetic clade 3C.2a, many subclades were formed, representing genetic diversification. Antigenic analysis revealed that almost all strains tested were antigenically different from cell culture-propagated A/Kansas/14/2017 (clade 3C.3a), the vaccine strain for the 2019/20 season (demonstrated 8-fold or greater reduction in virus neutralization (VN) titer than homologous VN titer of A/Kansas/14/2017 virus).
Only two domestic strains of B/Yamagata lineage were analyzed and they were antigenically similar to B/Phuket/3073/2013 virus (the vaccine strain for the 2019/20 season). Although all of the B/Victoria lineage strains belonged to genetic clade 1A of the HA gene tree, 99% of the domestic strains belonged to genetic clade 1A.3, which has three amino acid deletions in HA. The antigenic analysis also revealed that more than 90% of isolates in the 2019/20 season reacted well with ferret antiserum against B/Colorado/06/2017 virus (WHO recommended vaccine strain for the 2019/20 season), which has two amino acid deletions in HA.
Resistance to antivirals among 2019/20 isolates: Regarding neuraminidase inhibitors (oseltamivir, peramivir, zanamivir, and laninamivir), among the 2,489 domestic isolates of A/H1pdm09, 40 (1.6%) were resistant to oseltamivir/peramivir. Twenty-eight of these strains were isolated from patients who had not received these drugs. Regarding a cap-dependent endonuclease inhibitor (Baloxavir), among the 831 domestic isolates of A/H1pdm09, 1 (0.12%) Baloxavir-resistant mutant was detected from a patient who had not received the drug. As for A/H3 and B, all domestic isolates analyzed and isolates from 5 Asian countries were susceptible to NA inhibitors and Baloxavir (see p.200 of this issue).
Seroprevalence among the Japanese population prior to the 2019/20 season: Based on seroprevalence surveys conducted under the National Epidemiological Surveillance of Vaccine-Preventable Diseases program, the seroprevalence levels were measured from 5,446 serum samples collected between July and September 2019 (see p.201 of this issue). The age groups with the highest seroprevalence (measured as HI antibody-positive, titer ≥1:40) were 5-24-year-olds (57-60%) for A/Brisbane/02/2018 [A(H1N1)pdm09], 5-19-year-olds (53%) for A/Kansas/14/2017 [A(H3N2)], and 15-39-year-olds (60-70%) for B/Phuket/3073/2013 (B/Yamagata lineage). Compared to other age groups, seroprevalence for B/Maryland/15/2016 (B/Victoria lineage) was highest in 40-49-year-olds (51-52%).
Influenza vaccine: The tetravalent vaccine for the 2019/20 season contained antigens of two type A and two type B strains. Approximately 29,640,000 vial-equivalent doses (estimated on the assumption that 1 vial contained 1 mL) were produced, of which an estimated 28,250,000 vials were used.
For the 2020/21 season, A/Guangdong-Maonan/SWL1536/2019 (CNIC-1909) was selected as the A/H1 strain and A/Hong Kong/2671/2019 (NIB-121) as the A/H3 strain. As for the B/Yamagata strain, B/Phuket/3073/2013 was selected as in the 2019/20 season and B/Victoria/705/2018 (BVR-11) was selected for the B/Victoria strain (https://www.mhlw.go.jp/stf/shingi2/0000203023_00005.html). Active vaccination continues to be recommended.
A multicenter case-control study was performed on the effectiveness of two doses of the influenza vaccine, and for the 2018/19 season among children under three years of age, although not statistically significant, effectiveness against symptomatic influenza was observed (see p.204 of this issue).
Human infection with avian and swine influenza virus: From 2003 through March 2019, 861 confirmed human cases (455 fatal cases) of highly pathogenic avian influenza A(H5N1) virus infection were reported from 17 countries. Since October 2019, five countries, including China, reported cases of A(H5N1), four countries reported cases of A(H5N6), and 14 countries, including Japan, reported cases of A(H5N8). Since 2013, numerous human infection cases of avian influenza A(H7N9) have been reported in China, with a total of 1,568 cases (including 616 deaths). During the fifth epidemic from October 2016 to September 2017, A(H7N9) viruses that were highly pathogenic to poultry also emerged and human infection cases were reported, but no human infection case has been reported since the last case in March 2019. Human cases of avian influenza A(H9N2) virus infection have been sporadically reported from China, Egypt, Bangladesh, India, Senegal, and Oman.
Swine may play the role of an intermediate “mixing vessel” host, generating reassortments of avian and mammalian influenza viruses. Human cases of A(H3N2) variant (v), A(H1N1)v, and A(H1N2)v virus infections associated with exposure at agricultural fairs in the US have been reported; other swine influenza cases from Germany, China, and Brazil have also been reported. Swine influenza has also been detected in Japan and caution is needed (see p.206 of this issue).
Conclusion: The 2019/20 season saw an unprecedented decline in the reported number of influenza cases since the beginning of 2020. Care in interpretation is required, as this decrease may have been not only due to changes in individual-level behavior change or public health response measures, but also due to such aspects as refraining from visiting medical facilities, potentially affecting influenza activity and its relevant indicators. For the 2020/21 season, with the COVID-19 epidemic expected to continue, the impact of COVID-19 on influenza occurrence or virus detection is unclear--public health measures, including influenza vaccines, are being urgently put in place. At the same time, comprehensive monitoring through routine activities--such as the isolation/detection of influenza viruses, analyses of the antigenic and genetic properties of circulating strains, vigilance against the emergence of drug-resistant strains, and monitoring of population seroprevalence levels--will be ever more important. Further considerations on specific approaches will be necessary.
Note: For IASR reporting, influenza nomenclature is based on the virus information available. Influenza viruses are classified by type, subtype, and strain based on hemagglutination (HA), neuraminidase (NA), and other information:
- When both HA and NA typing have been performed, names are listed fully [e.g., A(H1N1)pdm09, A(H3N2), A(H5N1)].
- When NA typing has not been performed, only HA information is listed (e.g., A/H1pdm09, A/H3).
- The strain name is represented by the location of isolation; in the case of isolation in Japan, the location is written in Japanese kanji, and in the case of isolation in foreign countries, in English.
- To distinguish swine influenza viruses recovered from humans from seasonal influenza viruses, the term “variant virus” is used with a letter “v” added after the subtype name [e.g., A(H3N2)v].