

Influenza 2018/19 season, Japan
(IASR Vol. 40 p177-179: November, 2019)
The 2018/19 influenza season (from week 36 in September 2018 to week 35 in August 2019) was characterized by the predominance of the influenza A/H1pdm09 subtype in the beginning, followed by the AH3 subtype and B/Victoria lineage from week 10 in 2019.
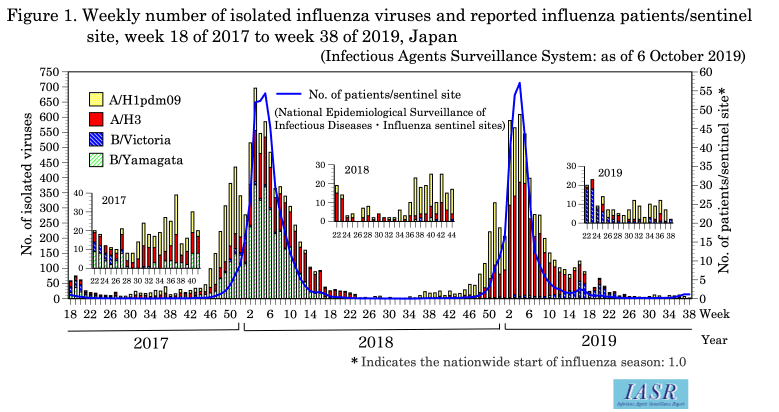
Epidemiology of the 2018/19 influenza season: Under the National Epidemiological Surveillance of Infectious Diseases (NESID) system, approximately 5,000 influenza sentinel sites (approximately 3,000 pediatric and 2,000 internal medicine facilities) report patients diagnosed as influenza on a weekly basis. The number of patients reported per sentinel exceeded 1.0 (the usual indicator of the nationwide start of influenza season) in week 49 of 2018 (Fig. 1 & https://www.niid.go.jp/niid/en/10/2096-weeklygraph/2572-trend-week-e.html) and exceeded 10.0 (indicator for the alert level used at the public health center level) in all 47 prefectures by week 2 of 2019. The peak was in week 4 of 2019 with 57.09 patients/sentinel, which was the highest since April 1999 when the Infectious Diseases Control Law was implemented.
{kind=link}
Based on the reported sentinel surveillance data, the estimated number of influenza patients who visited medical facilities between week 36 of 2018 and week 17 of 2019 was approximately 12,000,000. From the 2018/19 season, the method of estimation has been improved and is expected to be approximately 0.66-times the previous estimate. According to hospitalized influenza patient surveillance (data from approximately 500 designated sentinel hospitals with ≥300 beds), the number of hospitalized influenza patients (between week 36 of 2018 and week 17 of 2019) was 20,389. Among patients diagnosed as “acute encephalitis (including encephalopathy)” (a category V notifiable infectious disease), the causative pathogen was reported to be influenza virus in 223 (between week 36 of 2018 and week 17 of 2019). Approximately 3,400 excess deaths were observed nationwide in the 2018/19 season, which was estimated to be similar to the average year (see p. 192 of this issue).
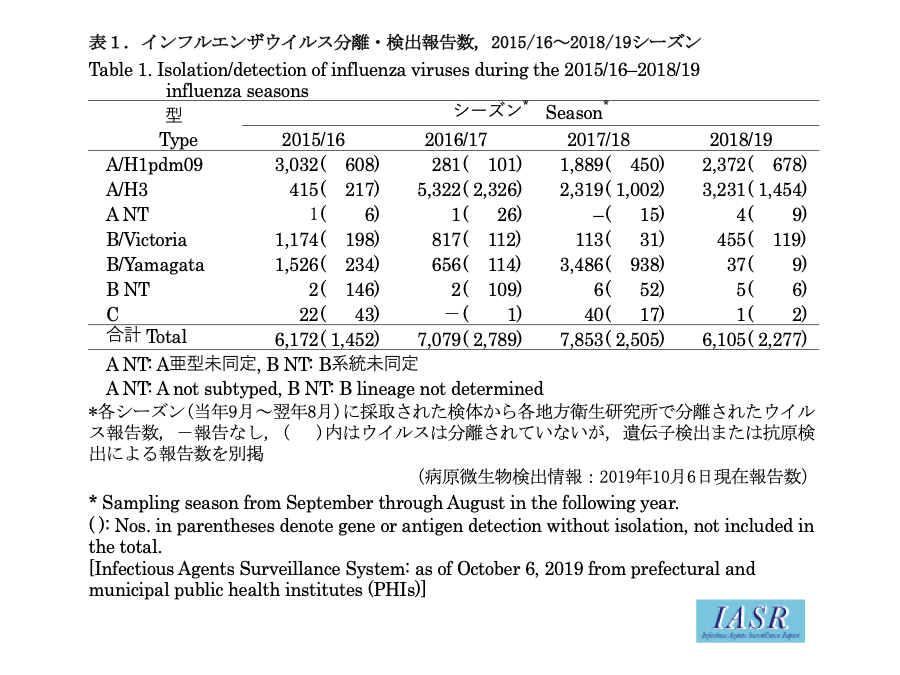
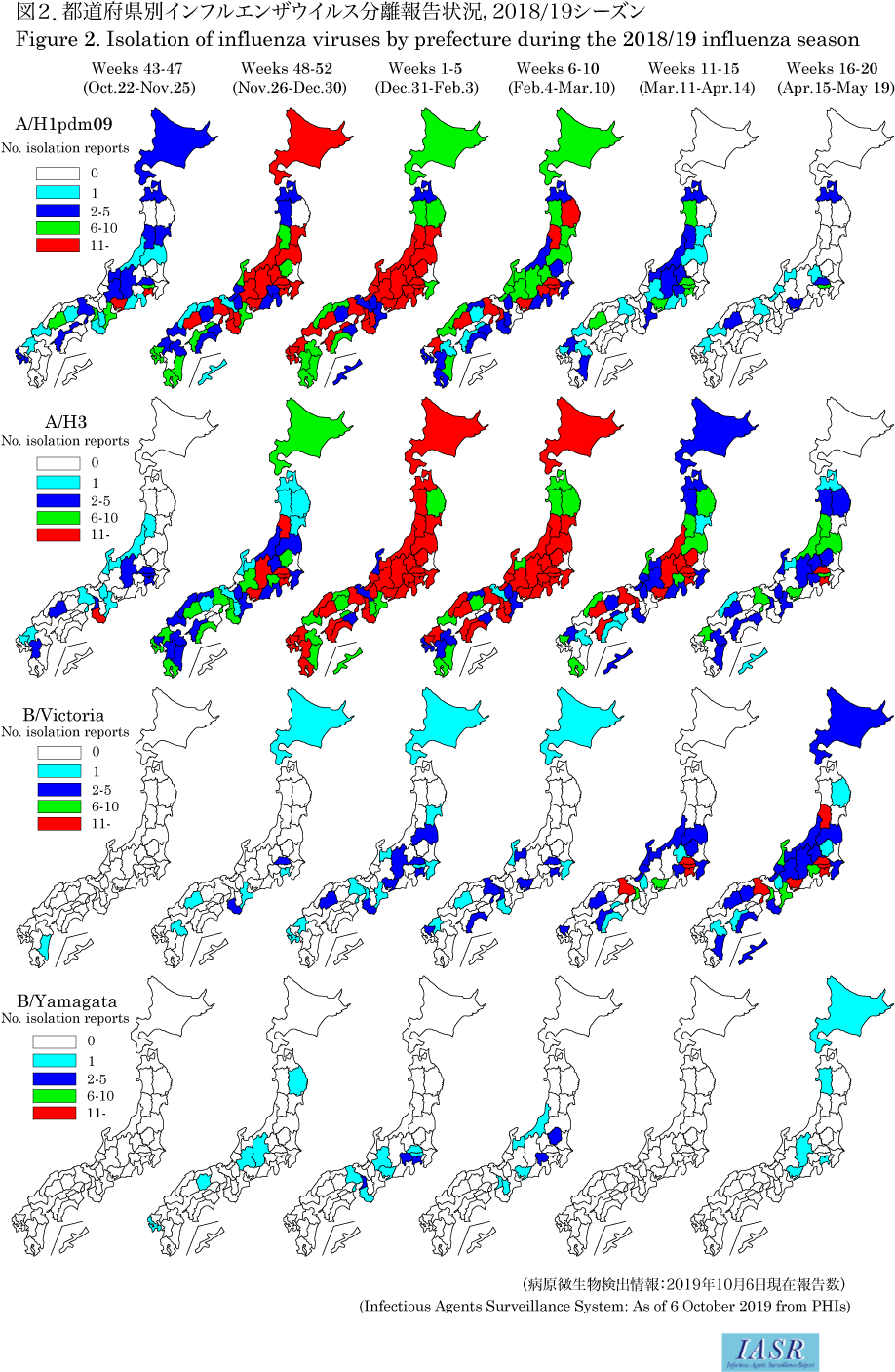
Isolation/detection of influenza virus in the 2018/19 season: Prefectural and municipal public health institutes (PHIs) reported the isolation/detection of 8,382 influenza viruses (6,105 isolates and 2,277 detections without isolation) (Table. 1). Among them, 7,704 were reported from the influenza sentinel sites and 678 from non-sentinel sites (Table 2). Influenza type A/H3 comprised 56%, A/H1pdm09 comprised 36%, and B comprised 8% (Yamagata lineage 7%, Victoria lineage 91%, and unknown lineage 2%) (Table 2). Although isolations of A/H1pdm09 began increasing from week 48 of 2018, it was surpassed by that of A/H3 from week 2 of 2019. Isolation of influenza type B began increasing from week 10 of 2019 and surpassed that of type A from week 17 of 2019 (Fig. 1 and Fig. 2). In the early summer of 2019, more A/H1pdm09 and A/H3 were found in Okinawa than in previous years (Fig. 2 and see p. 186 of this issue). Overseas, there were epidemics mainly of type A, whereas type B epidemics were less frequent.
{kind=link}
{kind=link}
{kind=link}
Genetic and antigenic characteristics of 2018/19 isolates: The National Institute of Infectious Diseases (NIID) conducts genetic and antigenic analyses of isolates from Japan and other Asian countries. For antigenic analysis, post-infection ferret antisera were used (see p. 180 of this issue). The hemagglutinin (HA) genes of all A/H1pdm09 isolates analyzed belonged to genetic clade 6B.1A, and the majority of viruses possessed a 183P substitution in HA. Antigenicity analysis demonstrated a lower reactivity of the viruses with vaccinated human serum. Although A/H3 is broadly divided into two groups, 3C.2a and 3C.3a, in the HA gene tree, all analyzed strains in the 2018/19 season belonged to 3C.2a. More than 90% of viruses tested reacted well with ferret antiserum against cell culture-propagated A/Singapore/ INFIMH-16-0019/2016 virus (the vaccine strain for the 2018/19 season), but hardly with the ferret antiserum against the egg-propagated virus, which had undergone antigenic mutation due to egg adaptation. The HA genes of all B/Yamagata lineage isolates tested belonged to genetic clade 3, and more than 90% of isolates tested were antigenically similar to B/Phuket/3073/2013 virus (the vaccine strain for the 2018/19 season). The HA genes of all B/Victoria lineage strains tested belonged to genetic clade 1A and 74% of isolates possessed 3 amino acid deletions in HA. Approximately 70% of viruses tested reacted well with ferret antiserum against B/Colorado/06/2017 virus (WHO recommended vaccine strain for the 2018/19 season), which has 2 amino acid deletions in HA.
Resistance to antivirals among 2018/19 isolates: Among the 2,163 domestic isolates of A/H1pdm09, 21 (1.0%) isolates were found to be resistant to oseltamivir/peramivir. Four of the strains were isolated from patients who had not been treated using the drug. Of the 331 A/H1pdm09 isolates from Japan, 6 (1.8%) baloxavir-resistant mutants were detected from patients treated using the drug. With the exception of one B isolate that was resistant to peramivir, the 331 A/H3 and 161 B domestic isolates analyzed were susceptible to oseltamivir, peramivir, zanamivir, and laninamivir. Regarding baloxavir, among 356 A/H3 isolates from Japan, 34 (9.6%) were resistant to baloxavir. Of these, 5 came from patients who had not been treated using baloxavir. Baloxavir-resistant variants of type B were not detected (see p. 185 of this issue).
Seroprevalence among the Japanese population prior to the 2018/19 season: Based on seroprevalence surveys conducted under the National Epidemiological Surveillance of Vaccine-Preventable Diseases program, the seroprevalence levels were measured from 6,569 serum samples collected between July and September 2018 (see p. 188 of this issue). The age groups with the highest seroprevalence (measured as HI antibody-positive, titer ≥1:40) were 10-24 year olds (68-71%) for A/Singapore/GP1908/2015 [A(H1N1)pdm09], 5-19 year olds (71-82%) for A/Singapore/INFIMH-16-0019/2016[A(H3N2)], and 20-34 year olds (66-70%) for B/Phuket/3073/2013 (B/Yamagata lineage). The sero-positivity for B/Maryland/15/2016 (B/Victoria lineage) was less than 40% for all age groups, except for 35-54 year olds (41-57%).
Influenza vaccine: The tetravalent vaccine for the 2018/19 season contained antigens of two type A and two type B strains. Approximately 27,170,000 vial-equivalent doses (estimated on the assumption that 1 vial contained 1 mL) were produced in the 2018/19 season, of which an estimated 26,300,000 vials were used.
For the 2019/20 season, A/Brisbane/02/2018(IVR-190) and A/Kansas/14/2017(X-327) were newly selected as the A/H1 strain and A/H3 strain, respectively. Both the B/Yamagata strain and B/Victoria strain selected were B/Phuket/3073/2013 and B/Maryland/15/ 2016(NYMC BX-69A), respectively, as in the 2018/19 season (see p. 196 of this issue).
A multicenter case-control study was performed on the effectiveness of two doses of the influenza vaccine in children under 6 years of age, and significant disease prevention effects were observed in the five seasons from 2013/14 to 2017/18 (see p. 194 of this issue). Continued active vaccination is recommended.
Human infection with avian and swine influenza virus (see p. 190 of this issue): Since 2003, 861 confirmed human cases (455 fatal cases) of highly pathogenic avian influenza A(H5N1) virus infection have been reported from 17 countries, including the first case in Nepal in March 2019. There have been 24 reports of A (H5N6) virus from China since 2014, including one human infection case in both March and August 2019 (as of September 27, 2019). On the other hand, since 2013, China has reported a total of 1,568 human cases (616 fatal cases) of low pathogenic avian influenza A(H7N9) virus infection (as of September 27, 2019). During the fifth epidemic (from October 2016 to September 2017), A (H7N9) viruses mutated to be highly pathogenic to poultry emerged, and human infection cases were also reported. However, only three cases of human infection due to highly pathogenic viruses have been reported since October 2017, and no cases have been reported after April 2019. Human cases of avian influenza A(H9N2) virus infection have been sporadically reported from China, Egypt, Bangladesh, and Oman.
Regarding swine influenza viruses, human cases of A(H3N2) variant (v), A(H1N1)v, and A(H1N2)v virus infections, associated with exposure at agricultural fairs in the United States, have been reported.
Conclusion: The response to influenza will continue to require sustained, comprehensive monitoring. Important activities include monitoring of trends in the occurrence of influenza patients, isolation of influenza viruses, analysis of the antigenic and genetic properties of circulating strains, vigilance against the emergence of drug-resistant strains, and monitoring of seroprevalence levels.
Note: For IASR reporting, influenza nomenclature is based on the virus information available. Influenza viruses are classified by type, subtype, and strain based on hemagglutination (HA), neuraminidase (NA), and other information:
- When both HA and NA typing have been performed, names are listed fully [e.g., A(H1N1)pdm09, A(H3N2), A(H5N1)].
- When NA typing has not been performed, only HA information is listed (e.g., A/H1pdm09, A/H3).
- The strain name is represented by the location of isolation; in the case of isolation in Japan, the location is written in Japanese kanji, and in the case of isolation in foreign countries, in English.
- To distinguish swine influenza viruses recovered from humans from seasonal influenza viruses, the term “variant virus” is used with a letter “v” added after the subtype name [e.g., A(H3N2)v].