Jpn. J. Infect. Dis., 56, 123-126, 2003
To see a printable version of the article in the Adobe file format, click this [PDF] link.
Laboratory and Epidemiology Communications
Drug Resistance Genes Encoded in Integrons and in Extra-Integrons: Their Distribution and Lateral Transfer among Pathogenic Enterobacteriaceae including Enterohemorrhagic Escherichia coli and Salmonella enterica Serovars Typhimurium and Infantis
Kokichi Hamada*, Kahori Oshima and Hidetaka Tsuji
Infectious Disease Research Division, Hyogo Prefectural Institute of Public Health and Environmental Sciences, Kobe 652-0032
Communicated by Kazue Tabita
(Accepted July 2, 2003)
*Corresponding author: Mailing address: Infectious Disease Research Division, Hyogo Prefectural Institute of Public Health and Environmental Sciences, Arata-cho 2-1-29, Hyogo-ku, Kobe 652-0032, Japan. Fax: +81-78-531-7080
Salmonella enterica serovars Typhimurium and Infantis have been major causes of Salmonella infections in Japan during the past decades, though S. Enteritidis suddenly emerged in 1989 and continues to prevail (1). While rare in S. Enteritidis, multidrug resistance (MDR) is frequent among S. Typhimurium and S. Infantis (1). The drug resistance genes are transferred among these Salmonella spp. along with the class 1 integrons (1) present in transposons and conjugative plasmids (1,2). Class 1 integrons are predominant within (2,3) and outside (4,5) the family Enterobacteriaceae. In Japan, enterohemorrhagic Escherichia coli (EHEC), particularly O157, O26, and O111 serotypes, has prevailed since 1996. Though well documented for O157 and O111 serotypes, integron-mediated antibiotic resistance among O26 serotype has remained relatively unknown (3). Here, we present a systematic investigation of drug resistance genes carried by integrons (2-5) or extra-integrons (2,4,6) in S. Typhimurium, S. Infantis, and EHEC with special reference to their transferability.
The strains used in this study were collected by authors. Salmonella strains were those used previously (1) and one additional S. Infantis strain (Inf32). EHEC strains were 29 serotype O26 strains (including two verotoxin non-producers), one O111, and three O157 strains, all collected in 1996-2003. The O111 and O157 EHEC strains were chosen randomly from a small number of MDR strains (resistant to more than three drugs) found in approximately 550 strains collected in 1996 - 2002.
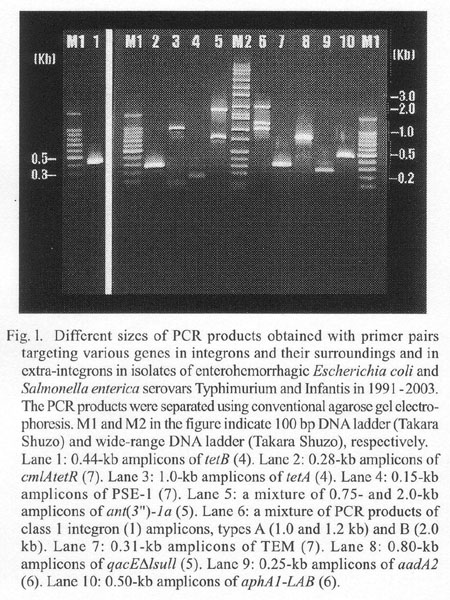
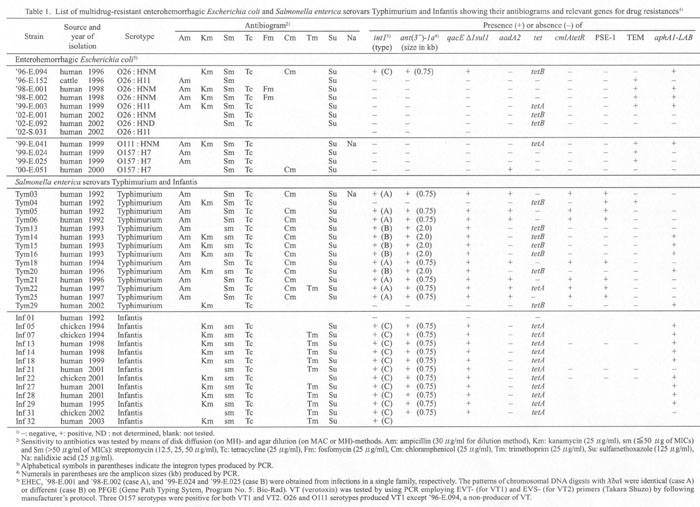
The strains were tested for sensitivities to ampicillin (Am), cefotaxime, kanamycin (Km), gentamicin, streptomycin (Sm), tetracycline (Tc), trimethoprim (Tm), ciplofloxacin, fosfomycin (Fm), chloramphenicol (Cm), sulphamethoxazole (Su), and nalidixic acid (Na). We used antibiotic disks (Becton Dickinson Microbiology Systems, Cockeysville, Md., USA) on Mueller-Hinton agar (MH) plates and agar dilutions on MacConkey (MAC) and/or MH agar plates (1). Table 1 shows the antibiograms of 14 MDR S. Typhimurium strains (among 22 strains tested), all the tested 12 MDR S. Infantis strains, and all the tested 11 MDR E. coli strains. A susceptible E. coli ('02-S.031) and a susceptible Salmonella (Inf01) were included for comparison. By means of polymerase chain reaction (PCR) (1), we searched for class 1 integrons, for ant (3")-1a and qacÂE1sul1 in close association with the 3'-conserved segment (3'-CS) of integron (2-5), and for drug resistance genes often located in integrons (2-5) or outside of integrons (2,4,6) . The primer pairs used for PCR and their PCR products are shown in Fig. 1. Primer concentration was 0.2 mM and Taq polymerase concentration 0.25 units/50 ml for all the genes except for tet genes (tetA, B, C, D, and E [4]). For PCR of tet, the primer concentration was 0.4 mM, and Taq polymerase concentration 0.5 units/50 ml.
Table 1 summarizes the characteristics of bacterial strains examined in the present and previous (1) reports.
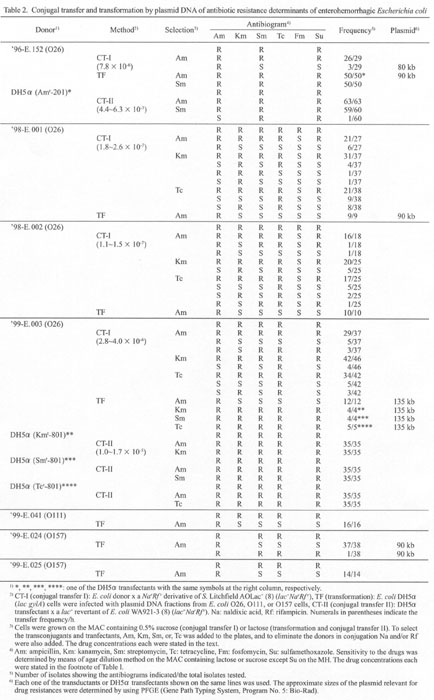
We examined 11 MDR strains of E. coli (Table
1) for transferability of drug resistance genes. Two protocols
were used. Conjugal transfer I (CT-I): Each of the E. coli
strains was conjugated with a rifampicin (Rf)-resistant derivative
of S. Litchfield AOLac+Nalr-01 (lac+Nar)
(1,8). The mating time was 4 h in liquid cultures at 37C. The
transconjugants were selected on sucrose (0.5%)-MAC plates containing
Am (30 mg/ml), Km (25 mg/ml),
Sm (25 and 50 mg/ml), or Tc (25 mg/ml). The donor strains were eliminated
by Na (25 mg/ml) and/or Rf (25 mg/ml). Conjugal transfer II (CT- II): In
order to know whether the resistance genes were present on transferable
plasmids or not, we conducted transformation followed by conjugal
transfer. For transformation, competent E. coli K12 DH5a (lac gylA) cells (Takara Shuzo,
Co., Ltd., Kyoto) were transfected with plasmid DNA fractions
prepared from the MDR strains. The transfectants were selected
on the lactose (0.5%)-MAC plates containing Am, Km, Sm, or Tc.
DH5a transfectants were crossed with
a lac+ revertant of E. coli K12 WA921-3
(lacNarRfr) (8). The MAC contained
Rf in order to eliminate the DH5a transfectants
used as donors. Table 2 shows CT-I and
CT-II data. The results are summarized as follows:
(1) Among 11 MDR EHEC, conjugal transfer (CT-1) was successful
for four strains, '96-E.152, '98-E.001, '98-E.002, and '99-E.003.
Segregation of drug resistance genes, especially Amr
from other resistant genes, was observed. This suggests Amr
and the other genes are located on a plasmid and the choromosome,
respectively (see also TF data in the table).
(2) CT-II was positive only for '96-E.152 and '99-E.003. In '96-E.152,
there was a rare segregation of resistance genes, AmsSmrSur
from AmrSmrSur, during
CT-II. For '99-E.003 no such segregation was observed.
Thus, four of 11 EHEC were able to transfer drug resistance genes through conjugation, and at least two ('96-E.152 and '99-E.003) of them harbored all the drug resistance genes also on the same plasmids.
REFERENCES
Go to JJID Homepage Go to JJID 56 (3) Contents
{kind=link}
{kind=link}
{kind=link}