Jpn. J. Infect. Dis., 56, 139-141, 2003
To see a printable version of the article in the Adobe file format, click this [PDF] link.
Laboratory and Epidemiology Communications
SARS Coronavirus-Infected Cells in Lung Detected by New In Situ Hybridization Technique
Noriko Nakajima, Yasuko Asahi-Ozaki, Noriyo Nagata, Yuko Sato, Florencio Dizon1, Fem J. Paladin1, Remigio M. Olveda1, Takato Odagiri2, Masato Tashiro2 and Tetsutaro Sata*
Department of Pathology and 2Department of Virology III, National Institute of Infectious Diseases, Tokyo 162-8640 and 1Research Institute for Tropical Medicine, Muntiniupa City, Philippines
Communicated by Tetsutaro Sata
(Accepted August 1, 2003)
*Corresponding author: Mailing address: Department of Pathology, National Institute of Infectious Diseases, Toyama 1-23-1, Shinjuku-ku, Tokyo 162-8640, Japan. Tel: +81-3-5285-1111, Fax: +81-3-5285-1189, E-mail: tsata@nih.go.jp
Severe acute respiratory syndrome (SARS) is an emerging infectious disease appearing in early 2003, resulting in 8,447 probable cases and 811 deaths around the world as of the end of June, 2003. Immediately after the SARS coronavirus (SARS-CoV) was isolated as an etiological agent, the complete sequences of the single-stranded positive sense RNA virus were deposited in databases and published (1). Although the lung pathology of SARS has been recently reported (2), the site of the virus replication and its role in the pathogenesis of SARS-CoV-associated lung disease remain uncertain because of the lack of a specific antibody to detect the virus antigen by immunohistochemistry in tissue sections. Recently, we developed a new in situ hybridization (ISH) technique, the in situ hybridization AT tailing coupled with catalized signal amplification (ISH-AT-CSA) method, which is superior in terms of both sensitivity and specificity to conventional ISH (3). This method consists of hybridization with an oligonucleotide probe which has an oligo d(A-T) tag on the 3' end, followed by elongation of d(A-T) by DTth DNA polymerase in the presence of biotin-16-dUTPs. In the ISH-AT-CSA method, only an oligonucleotide probe complementary to the target sequence needs to be prepared for visualizing gene expression at the cellular level in tissues. We obtained a formalin-fixed lung tissue of an autopsied SARS patient from the Philippines, a 46-year-old woman who fulfilled the case definition and died of respiratory failure. To detect the SARS-CoV genome and/or transcripts, ISH-AT-CSA using an oligonucleotide probe consisting of a 40-base complementary sequence of the nucleocapsid region of SARS-CoV and (AT)10 on the 3' side (antisense; 5'biotin-gat aag tca gcc atg ttc ccg aag gtg tga ctt cca tgc cat ata tat ata tat ata tat at-3' , sense; 5'biotin-ggc atg gaa gtc aca cct tcg gga aca tgg ctg act tat cat ata tat ata tat ata tat-3') was applied. Here we show first SARS-CoV replication occurring mainly in alveolar epithelial cells.
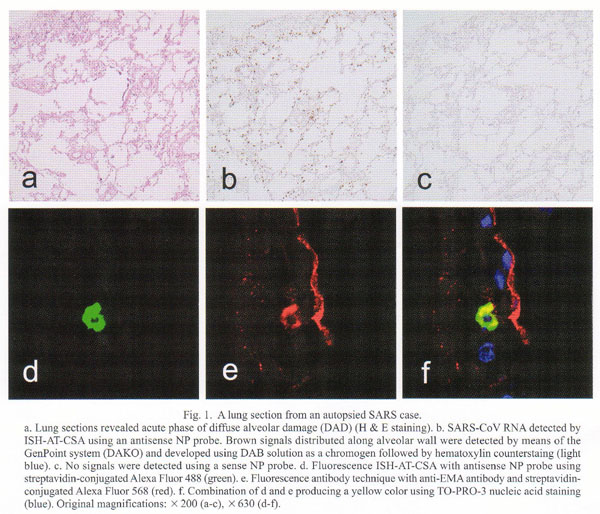
Formalin-fixed and paraffin-embedded lung sections revealed mild diffuse alveolar damage (DAD). Neither inclusion-bearing cells nor multinucleated giant cells were seen. Their histopathologic findings varied among lobules, suggesting a trans-bronchobronchiolar spread of the infection (Fig. 1a). They were divided roughly into three patterns: 1) mild inflammatory cell infiltration without exudation or hyaline membrane, 2) with small amount of exudation without hyaline membrane, and 3) with hyaline membrane, edema, and vascular congestion, which were totally consistent with acute phase of DAD. Immunohistochemistry for CD68 (PGM1, Dako, Kyoto) disclosed mild increase of macrophages distributed evenly in the section (data not shown). ISH-AT-CSA using an antisense NP probe could detect viral RNA in the cytoplasm of several cells (Fig. 1b). The sense NP probe could not detect any signals in the cells (Fig. 1c). In order to determine the phenotype of the virus-infected cells, confocal laser scanning microscopic studies by double staining with fluorescence ISH-AT-CSA and fluorescence antibody technique for CD68, epithelial membrane antigen (EMA), surfactant apoprotein A, or CD34 (Dako) were performed. The ISH-AT-CSA signals (Fig. 1d, green) and EMA signals (Fig. 1e, red) were merged and produced yellow signals (Fig. 1f), which confirmed that the virus-infected cells were alveolar epithelial cells. Other double fluorescence stainings showed that viral RNA was detected also in a few macrophages, and a few bronchiolar epithelial cells, but not in endothelial cells (data not shown). As control studies, lung specimens from autopsied patients with influenza, measles, varicella-zoster virus infection, and normal appearing lungs revealed no signals. Furthermore, ISH-AT-CSA by using unrelated probes or the omission of DTth DNA polymerase in the reaction mixture failed to detect signals. This localization of the SARS-CoV suggests that virus replication occurred mostly in alveolar epithelial cells, to a lesser extent in alveolar macrophages in the area of mild inflammatory cell infiltration without exudation, and rarely in the area of DAD lesions.
To confirm SARS-CoV infection in the lung tissue, we conducted RT-PCR assay and electron microscopic examination. A small amount of the fixed lung tissue was sampled. RNA was extracted by using Trizol (Invitrogen, Tokyo) according to the manufacturerŐs instructions. The RNA was dissolved in TE buffer, and heated at 70C for 30 min in order to enhance the reverse transcription reaction. Two sets of primers (BNIout and SAR1) specific to SARS-CoV provided by the World Health Organization (PCR primers for SARS developed by WHO Network Laboratories are available at http://www.who.int/csr/sars/primers/en/) were synthesized and utilized for both RT reaction and PCR amplification. The expected sized bands (190 and 121 bp, respectively) were amplified and purified for direct sequencing. The sequences obtained were those of SARS-Cov. The lung tissue was also subjected to electron microscopic examination. Epon-embedded thin sections showed the presence of viral particles, around 100 nm in diameter, in the cytoplasm of alveolar epithelial cells and macrophages.
In the early phase of a newly emerging virus-associated infectious disease, specific histological diagnosis and investigation of the virus-infected cells in the tissues by immunohistochemistry are impossible due to the lack of specific antibodies. Our new ISH technique requires only information regarding a part of the sequence. The ISH-AT-CSA method may provide a valuable tool for the histologic diagnosis and analysis of the pathogenesis of emerging infectious diseases. Although the examined number of SARS cases was limited in this report, we postulate that the virus spreads via a broncho-bronchial route and replicates mainly in the alveolar epithelium to cause DAD.
We thank the WHO Regional Office for the Western Pacific for providing the opportunity to examine lung tissue.
REFERENCES
Go to JJID Homepage Go to JJID 56 (3) Contents
{kind=link}