Jpn. J. Infect. Dis., 56, 216-218, 2003
To see a printable version of the article in the Adobe file format, click this [PDF] link.
Laboratory and Epidemiology Communications
Chromosomal Transferable Multidrug Resistance Genes of Salmonella enterica Serovar Infantis
Kokichi Hamada*, Kahori Oshima and Hidetaka Tsuji
Infectious Disease Research Division, Hyogo Prefectural Institute of Public Health and Environmental Sciences, Kobe 652-0032
(Accepted November 10, 2003)
Communicated by Kazue Tabita
*Corresponding author: Mailing address: Infectious Disease Research Division, Hyogo Prefectural Institute of Public Health and Environmental Sciences, Arata-cho 2-1-29, Hyogo-ku, Kobe 652-0032, Japan. Fax: +81-78-531-7080
In Japan, Salmonella enterica serovar Infantis, together with S. enterica serovar Typhimurium, has been one of the major causes of Salmonella infections during the past decades, though in 1989 S. enterica serovar Enteritidis suddenly emerged and its epidemic continues (1-3). While rare among S. Enteritidis (3), multidrug resistance (MDR) was frequent among S. Infantis and S. Typhimurium (1,2). MDR genes were transferable along with or without a class 1 integron among them (1,2) and also among enterohemorrhagic Escherichia coli (2). The present communication shows the chromosomal localization of MDR genes in S. Infantis.
Among 32 S. Infantis isolates (Inf01-Inf32) from 1992 - 2003 in Hyogo Prefecture, 23 isolates were implicated in three food poisoning outbreaks and 15 sporadic infections and were of human sources (see the legend to Fig. 1). The remaining nine isolates were from other than humans, one from causative food in an outbreak (Inf25), six from chickens/eggs (Inf03, Inf04, Inf05, Inf07, Inf22, and Inf31), and two from the environment (Inf06 and Inf30). They had no mutual epidemiological relations except between Inf08 and Inf10 from an epidemic in 1996, between Infl5 and Infl7 in an epidemic in 1999, and between Inf23 and Inf25 in an epidemic in 2001.
All the isolates were tested for drug sensitivities by the disk method (Becton Dickinson Microbiology Systems, Cockeysville, Md., USA) (1,2) and by the dilution method. For the dilution method, lactose (0.5%) - MacConkey agar (Lac-MAC) plates were used for testing sensitivities to kanamycin (Km, 25 mg/ml), streptomycin (Sm, 12.5, 25, and 50 mg/ml), and tetracycline (Tc, 25 mg/ml), and Mueller-Hinton (MH) plates for testing sensitivities to trimethoprim (Tm, 25 mg/ml) and sulfamethoxazole (Su, 125 mg/ml). Twelve isolates had MDR that could be classified into three groups. Eight isolates (Inf07, Infl3, Infl4, Infl8, Inf27, Inf28, Inf29, and Inf32) were resistant to Km, Sm, Tc, Tm, and Su, two (Inf05 and Inf22) to Km, Sm, Tc, and Su, and two (Inf21 and Inf31) to Sm, Tc, Tm, and Su (2). Among them Inf05, Inf07, Inf22, and Inf31 were from chickens/eggs and the remaining eight were from humans (2).
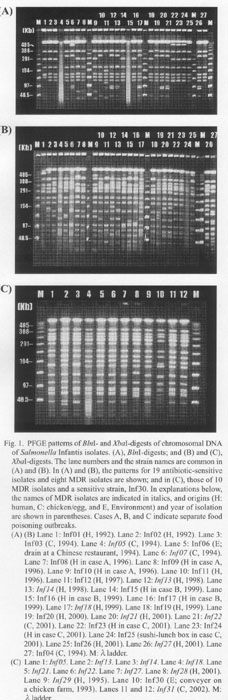
Figure 1 shows the results of pulsed-field gel electrophoresis (PFGE) (Gene Path Typing System, Program No. 5; Bio-Rad Laboratories, Hercules, Calif., USA) of BlnI- or XbaI-digests of chromosomal DNA. The patterns of all of the isolates digested with BlnI (Fig. 1A) were not sharply differentiated. The pattern was dissimilar among the XbaI-digests of the isolates (Fig. 1B), though less variable among those of the MDR isolates (Fig. 1B, 1anes 4, 6, 12, 13, 17, 20, 21, and 26; Fig. 1C), which may indicate a common genetic background of these MDR strains.
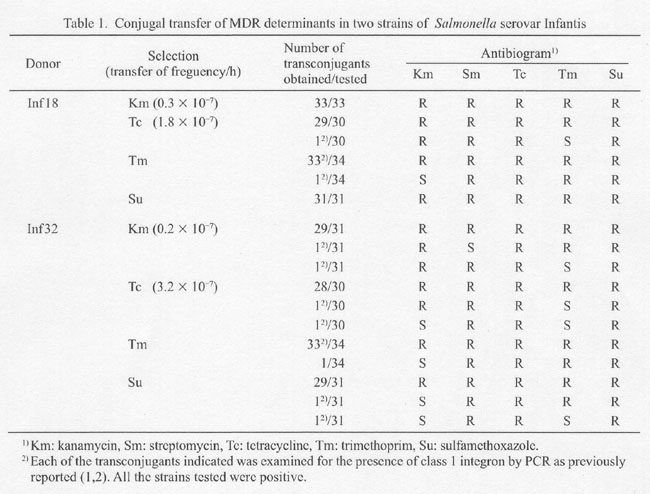
To locate drug resistance, we mated donor isolates Infl8 and Inf32, which were KmrSmrTCrTmrSur, with a recipient S. enterica serovar Litchfield AOLac+Nalr-01 (lac+Nar) (4) for 4 h in liquid cultures at 37Åé. The transconjugants were selected on Lac-MAC- (for Kmr or Tcr) or MH-plates (for Tmr or Sur). The donor was eliminated by nalidixic acid (Na) (25 mg/ml). Segregation of drug resistance genes during the conjugation was rare, however it was certainly observed (Table 1). Plasmids from nine MDR isolates failed to transfer resistance genes (data not shown). Thus, drug resistance genes were not on a plasmid but on the chromosome.
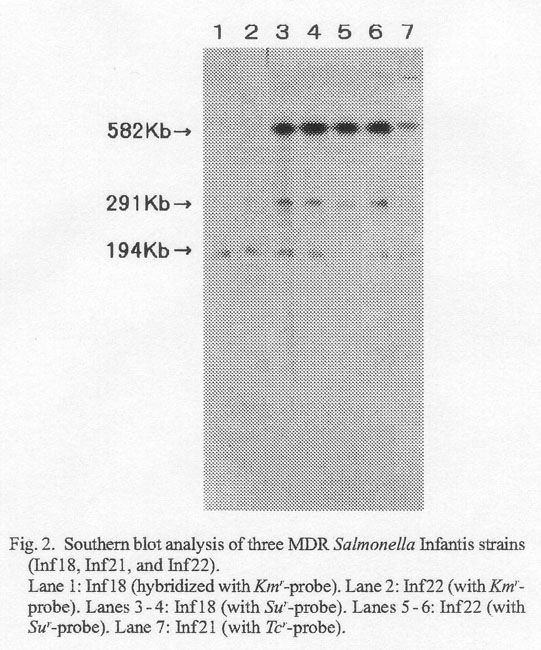
Southern blot analysis was conducted on Inf18, Inf21, and Inf22 by using aphA1-LAB as Kmr-probe, TetA as Tcr-probe, and qacED1sul1 as Sur-probe (2). The probes were prepared by polymerase chain reaction, which incorporated digoxigenin-11-dUTP (Boehringer GmbH, Mannheim, Germany) in the reaction mixture, using template Infl8 (KmrSmrTcrTmrSur) DNA and appropriate primer pairs (2). Genomic DNA of Inf18, Inf21 (SmrTcrTmrSur), and Inf22 (KmrSmrTcrSur) were digested with XbaI, run on PFGE (Gene Path Typing System, Program No. 5; Bio-Rad) and transferred to nylon membranes. Figure 2 shows the presence of Kmr and Sur genes on the chromosomes of Inf18 and Inf22 (lanes 1-6). As expected, chromosomal DNA of Inf21 hybridized with Tcr probe (lane 7) but not with Kmr-probe (data not shown).
REFERENCES
Go to JJID Homepage Go to JJID 56 (5,6) Contents
{kind=link}
{kind=link}
{kind=link}