Jpn. J. Infect. Dis., 54, 122-124, 2001
To see an article, click this [PDF] link.
Laboratory and Epidemiology Communications
Sensitive Detection of Cryptosporidium Oocysts in Environmental Water Samples by Reverse Transcription-PCR
Tatsuya Karasudani*, Satomi Aoki, Junko Takeuchi, Masaaki Okuyama, Mitsuaki Oseto, Shigemi Matsuura, Tadao Asai and Hiroo Inouye
Ehime Prefectural Institute of Public Health and Environmental
Science,
Sanban-cho 8-234, Matsuyama, Ehime 790-0003
Communicated by Hiroo Inouye
(Accepted July 24, 2001)
Cryptosporidium is present in the environment as an oocyst
andü@causes outbreaks through contamination of the water supply.
With a reverse transcription-PCR (RT-PCR) amplifying heat shock
protein 70 (hsp70) gene, we were able to detect two different
types of viable Cryptosporidium oocysts from the environment.
Water specimens were filtered under positive pressure through a 3-mm polycarbonate filter (Nuclepore filter, Whatman, Maidstone, England). The trapped materials were detached from the filter by sonication, and concentrated by centrifugation. The concentrates were subjected to mRNA extraction and RT-PCR amplification in the manner described by Kaucner et al. (1). C. parvum HNJ1 supplied by Dr. Kuroki, Kanagawa Prefectural Institute of Public Health, was used as a positive control.
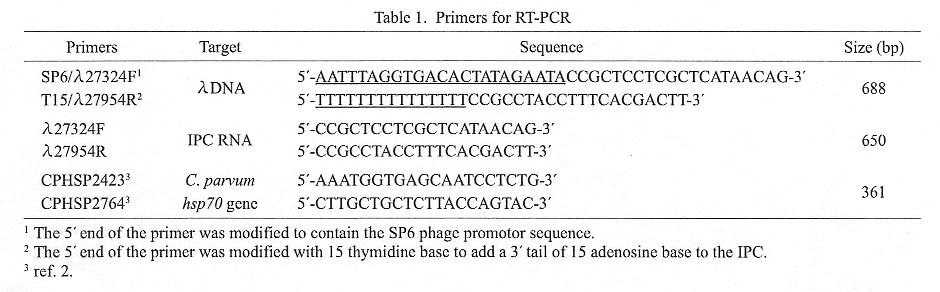
The concentrated specimens were suspended in InstaGene matrix (Bio-Rad, Hercules, Calif., USA), and heated for 20 min at 45üÄ for induction of hsp70 gene expression. After freeze-thaw, mRNA was extracted from the samples using the Dynabeads mRNA DIRECT kit (Dynal, Oslo, Norway), the hsp70 gene was RT-PCR amplified. To monitor the sensitivity of the RT-PCR amplification, a poly A-tailed RNA was included in the reaction as an internal positive control (IPC RNA). The IPC RNA was a transcript from an artificial gene, which consisted of an SP6 promoter and nt 27, 324-27, 974 of the ā╔-phage genome, followed by 15 Ts on the complementary strand. The artificial gene (688 bp in size) was first PCR-amplified, and then used for transcription of IPC RNA by the Competitive RNA Transcription kit (TAKARA, Kyoto). Primers used for RT-PCR are listed in Table 1. SP6/ā╔27324F and T15/ā╔27954R were the primers used for PCR amplification of the 688-bp template DNA, andā╔27324F and ā╔27954R for RT-PCR amplification of IPC RNA. CPHSP2423 and CPHSP2764 were used as primers for amplification of the C. parvum hsp70 gene.
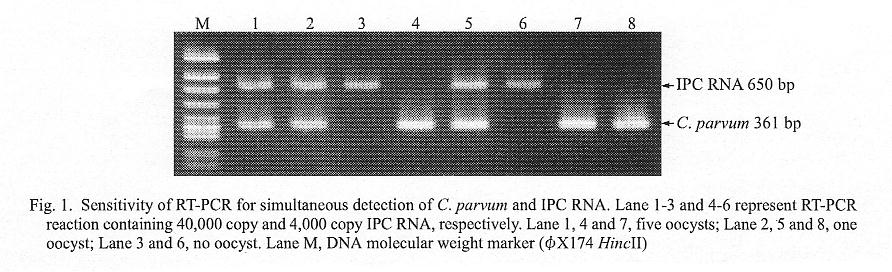
The effects of inclusion of IPC RNA and its PCR primers on the specific Cryptosporidium hsp70 gene amplification was examined. As shown in Fig. 1, the presence of 4,000 copies of IPC RNA had no effect (compare lanes 4 and 5 with lanes 7 and 8); addition of 10-fold more copies, i.e., 40,000 copies of IPC RNA, tended to decrease the density of the specific band (lanes 1 and 2), but even under these conditions, the sensitivity was sufficient for detecting a single viable C. parvum oocyst.
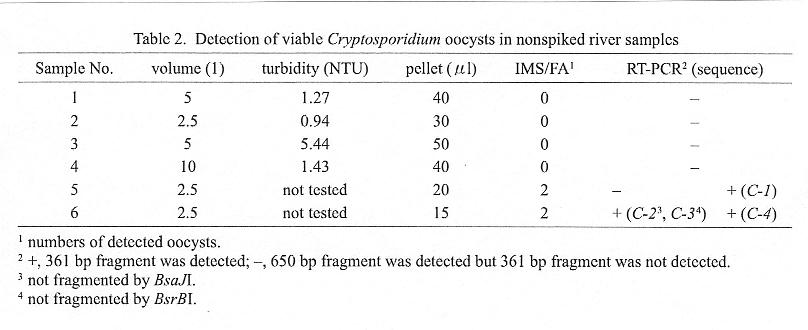
Using the above RT-PCR and immunomagnetic separation and immunofluorescence assay (IMS/FA), we attempted to detect Cryptosporidium from river-water specimens obtained at different locations (Table 2). IMS/FA detected two C. parvum-like oocysts each from the No. 5 and No. 6 specimens. In the RT-PCR assays performed twice for each sample, No. 5 was positive in one trial and No. 6 in both trials. The RT-PCR products were recovered from the gel, and their sequences were determined by the Gene Rapid (Amersham Pharmacia Biotech, Piscataway, N.J., USA) dye terminator method. One of the two No. 6 specimens was found to be a mixture of at least two variants, as variations of two nucleotides were observed at 15 locations in the amplified 361-nt fragment. As only one variant was cut by BsaJI and BsrBI, the sequence of the uncut fragment was determined in order to deduce the sequences of the two variants.
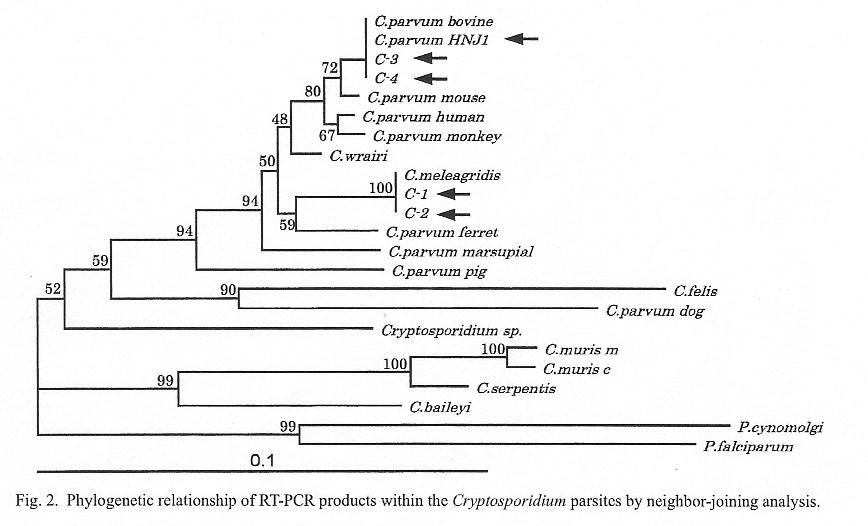
In total, four sequences were obtained from this study. The phylogenetic tree was constructed from these sequences together with published Cryptosporidium hsp70 sequences (3). The sequences of C-3 and C-4 were identical to those of the C. parvum bovine genotype, and those of C-1 and C-2 were identical to those of C. meleagridis (Fig. 2), i.e., specimen No. 5 contained C. meleagridis, and specimen No. 6 the C. parvum bovine genotype and C. meleagridis. C. meleagridis has been considered to be a parasite of turkeys, but its infection to humans has recently been confirmed (4). It has been suggested that bird discharge may be a source of Cryptosporidium contamination of the water (5). Our results demonstrate that RT-PCR may be a useful tool for molecular epidemiology of Cryptosporidium.
REFERENCES
*Corresponding author: Fax: +81-89-947-1262, E-mail: karasudani-tatsuya@pref.ehime.jp
Go to JJID Homepage Go to JJID 54(3) Contents
{kind=link}
{kind=link}
{kind=link}
{kind=link}