|
国立感染症研究所インフルエンザウイルス研究センター第一室
全国地方衛生研究所 |
| 流行株抗原性解析 |
| 国立感染症研究所(感染研)では、国内で流行するインフルエンザウイルスの性状を把握し、インフルエンザ対策およびワクチン株選定に役立てるため、全国地方衛生研究所(地研)で分離・同定されたウイルス株総数の約10%を無作為に抽出し、解析を行っている。 |
| 流行株とワクチン株の抗原性を比較する目的で、フェレット感染血清を用いた赤血球凝集阻止(HI)試験または中和試験による抗原性解析を実施した。 |
|
現行の季節性インフルエンザワクチンは、ワクチン原株として選ばれたウイルスを鶏卵で継代して製造している。そのため、継代の間に、ウイルスが鶏卵に馴化することでアミノ酸置換が起こり、抗原性が変化(抗原変異)することがある。その結果、流行株とワクチン製造株の抗原性が一致しなくなる場合があり、世界的に問題となっている。 |
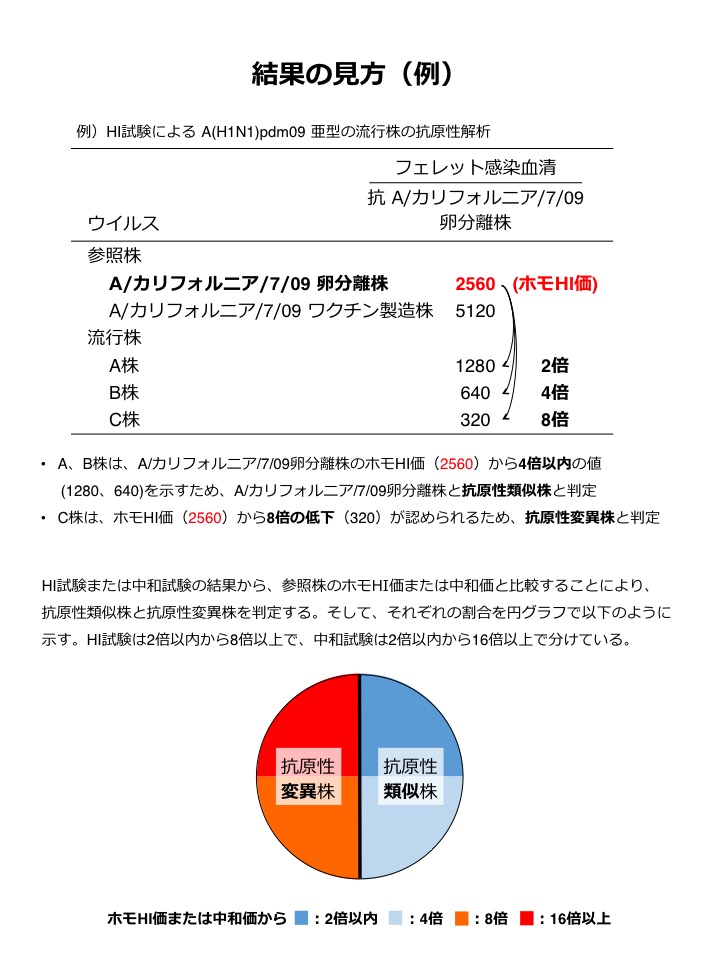
| 抗原性解析試験:結果の見方 |
{kind=link}
|
2017/2018シーズン抗原性解析結果(データ更新日:2018年2月19日)NEW |
|
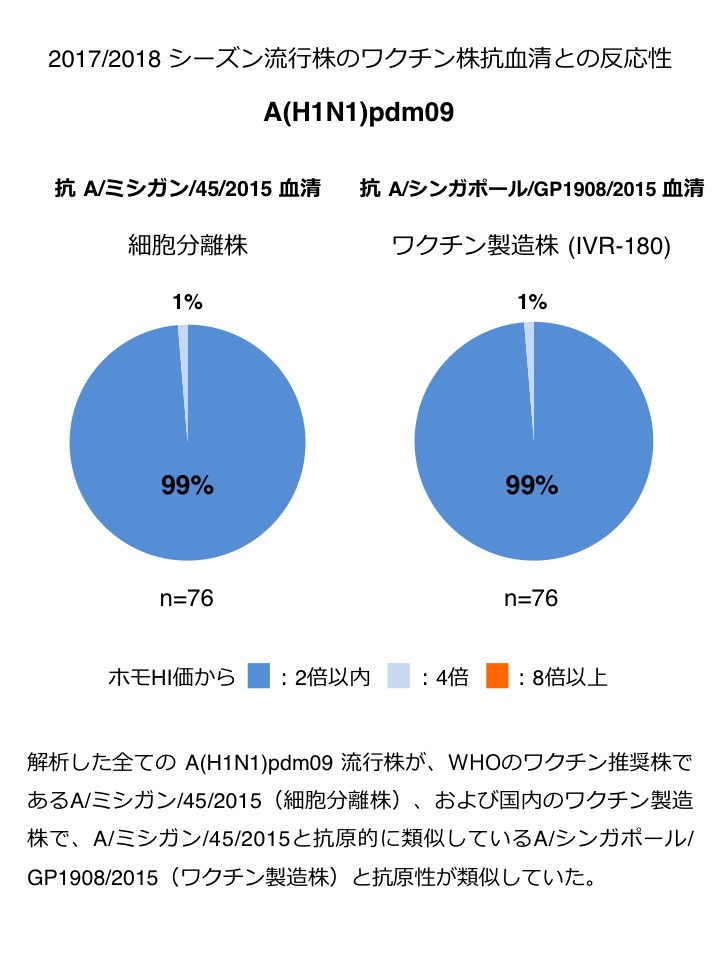
A(H1N1)pdm09 : 解析した全ての A(H1N1)pdm09 流行株が、WHOのワクチン推奨株であるA/ミシガン/45/2015(細胞分離株)、および国内のワクチン製造株で、A/ミシガン/45/2015と抗原的に類似しているA/シンガポール/GP1908/2015(ワクチン製造株)と抗原性が類似していた(図1)
|
{kind=link}
|
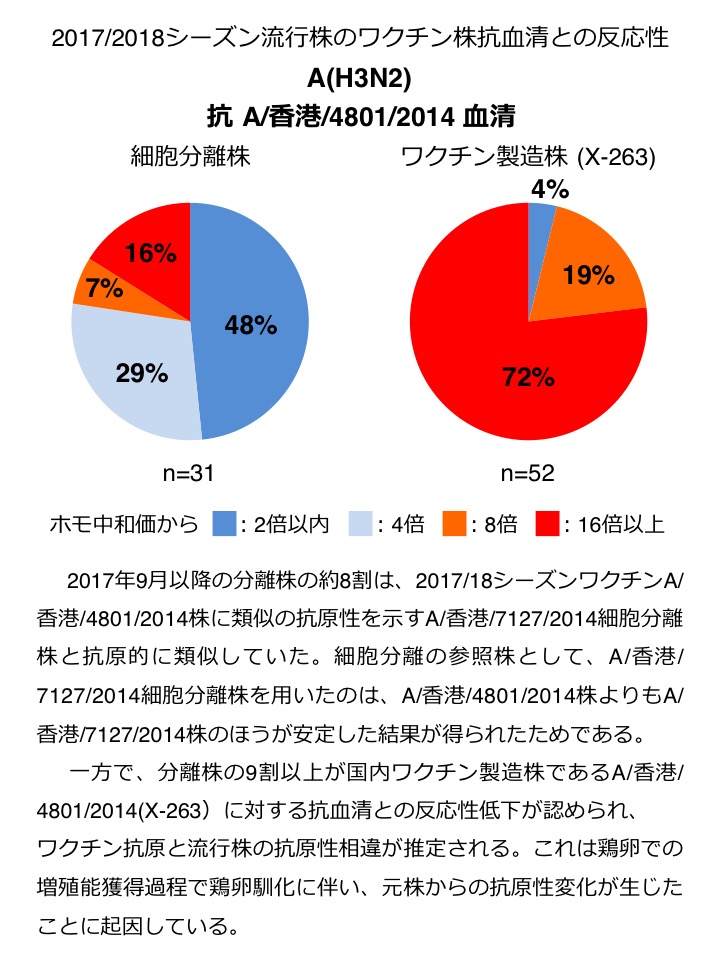
A(H3N2) : 2017年9月以降の分離株の約8割は、2017/18シーズンワクチンA/香港/4801/2014株に類似の抗原性を示すA/香港/7127/2014細胞分離株と抗原的に類似していた。細胞分離の参照株として、A/香港/7127/2014細胞分離株を用いたのは、A/香港/4801/2014株よりもA/香港/7127/2014株のほうが安定した結果が得られたためである。一方で、分離株の9割以上が国内ワクチン製造株であるA/香港/4801/2014(X-263)に対する抗血清との反応性低下が認められ、ワクチン抗原と流行株の抗原性相違が推定される。これは鶏卵での増殖能獲得過程で鶏卵馴化に伴い、元株からの抗原性変化が生じたことに起因している(図2)。
|
{kind=link}
|
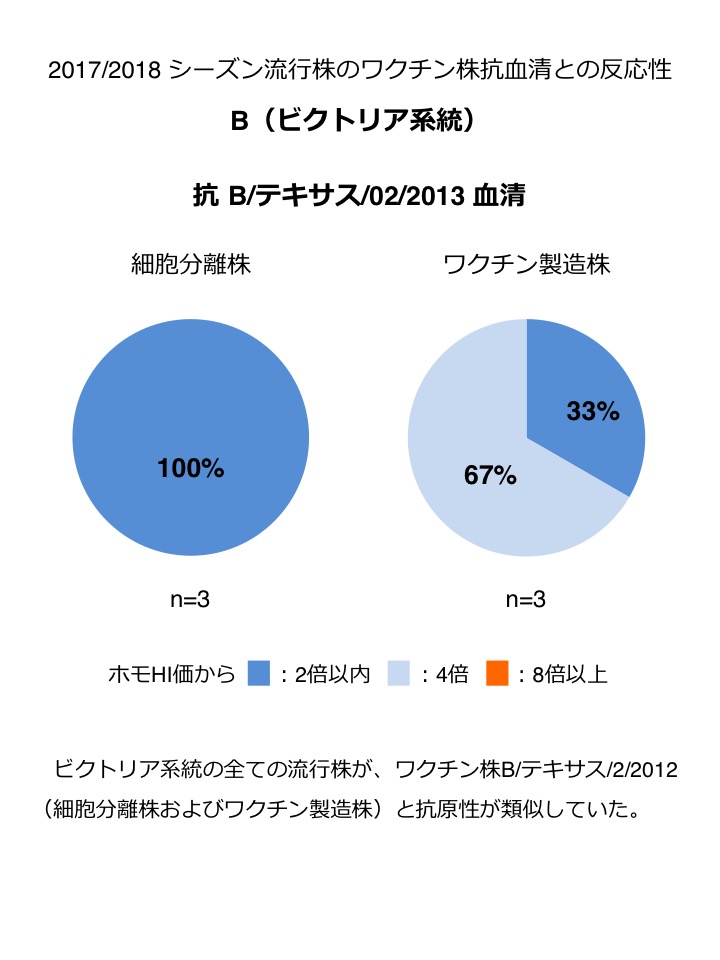
B(ビクトリア系統) : 解析数は少ないが、2017年9月以降の流行株は、ワクチン株B/テキサス/2/2012(細胞分離株およびワクチン製造株)と抗原性が類似していた(図3)。
|
{kind=link}
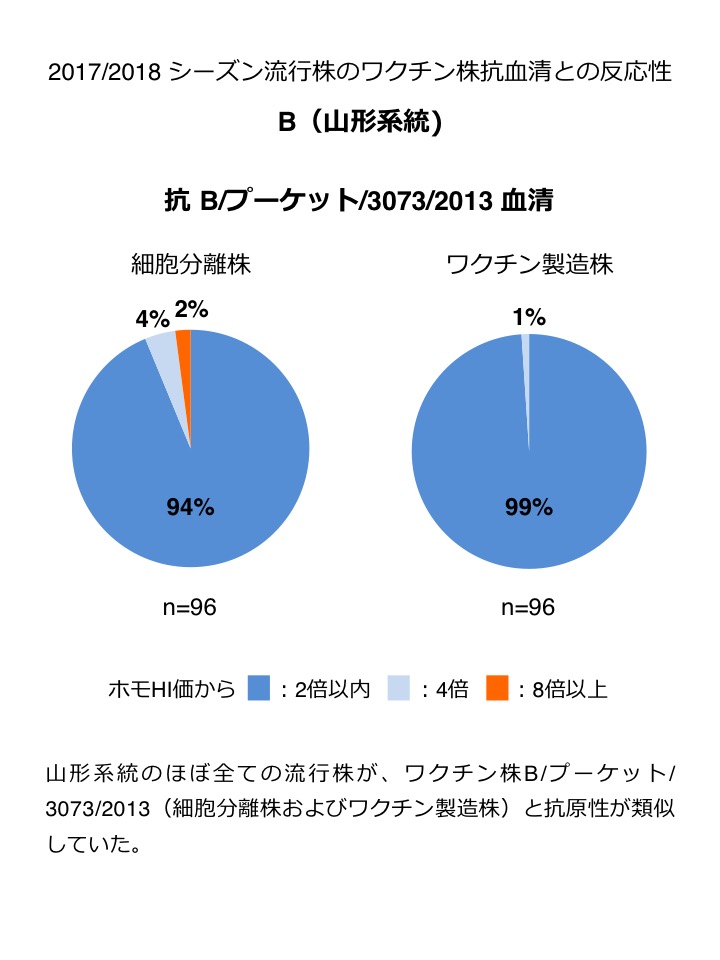
| B(山形系統) : 2017年9月以降で解析したほぼ全ての流行株は、ワクチン株 B/プーケット/3073/2013(細胞分離株およびワクチン製造株)と抗原性が類似していた(図4)。 |
{kind=link}
| 遺伝子系統樹 |
| 国立感染症研究所インフルエンザウイルス研究センター第一室が解析した季節性インフルエンザウイルスの遺伝子配列を用いて、HA遺伝子系統樹を作成した。国内外で流行しているウイルスと比較するため、各地方衛生研究所にて分離された株の遺伝子配列だけではなく、海外で分離された株の遺伝子配列も解析に加えている。なお、海外の研究機関で解析された遺伝子配列はインフルエンザウイルス遺伝子データベースGISAID(Global Initiative on Sharing All Influenza Data:http://platform.gisaid.org/epi3/frontend)から入手している。 |
| 2017/2018シーズン系統樹(データ更新日:2018年1月29日) |
|
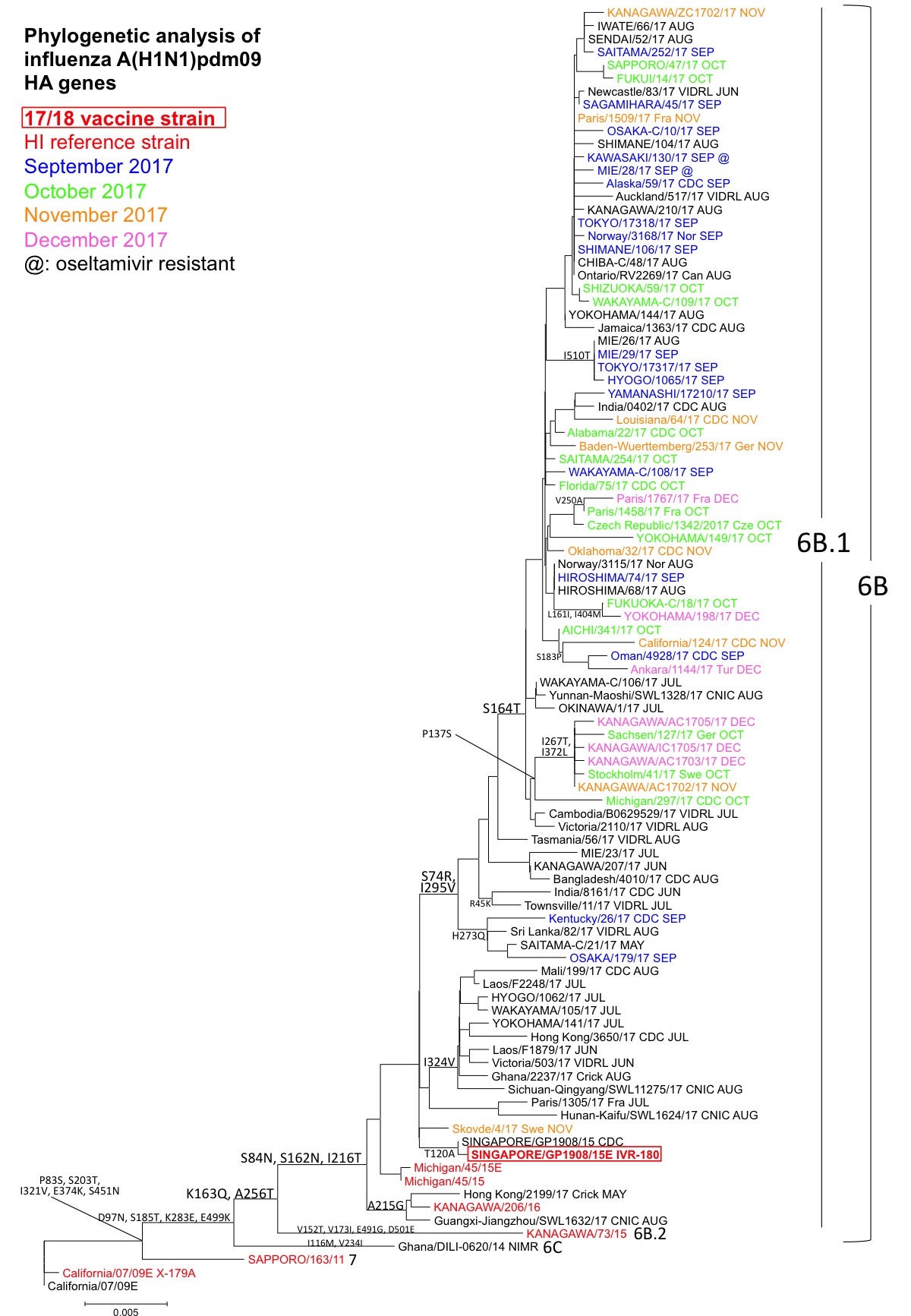
A(H1N1)pdm09:解析した株は全てHA遺伝子系統樹上のクレード6B.1(共通アミノ酸置換:S84N, S162N, I216T)に属していた(図1)。サブクレード6B.1内で集団はさらに分岐し、S74R, I295V群、I324V群を形成した。S74R, I295V群には、さらに、S164T群、H273Q群が形成されているが、S164T群に属するウイルスが多く検出されている。また、NAタンパク質にH275Y置換を有するオセルタミビル耐性株は散発的に検出されているが、耐性株の流行は確認されていない。
|
{kind=link}
|
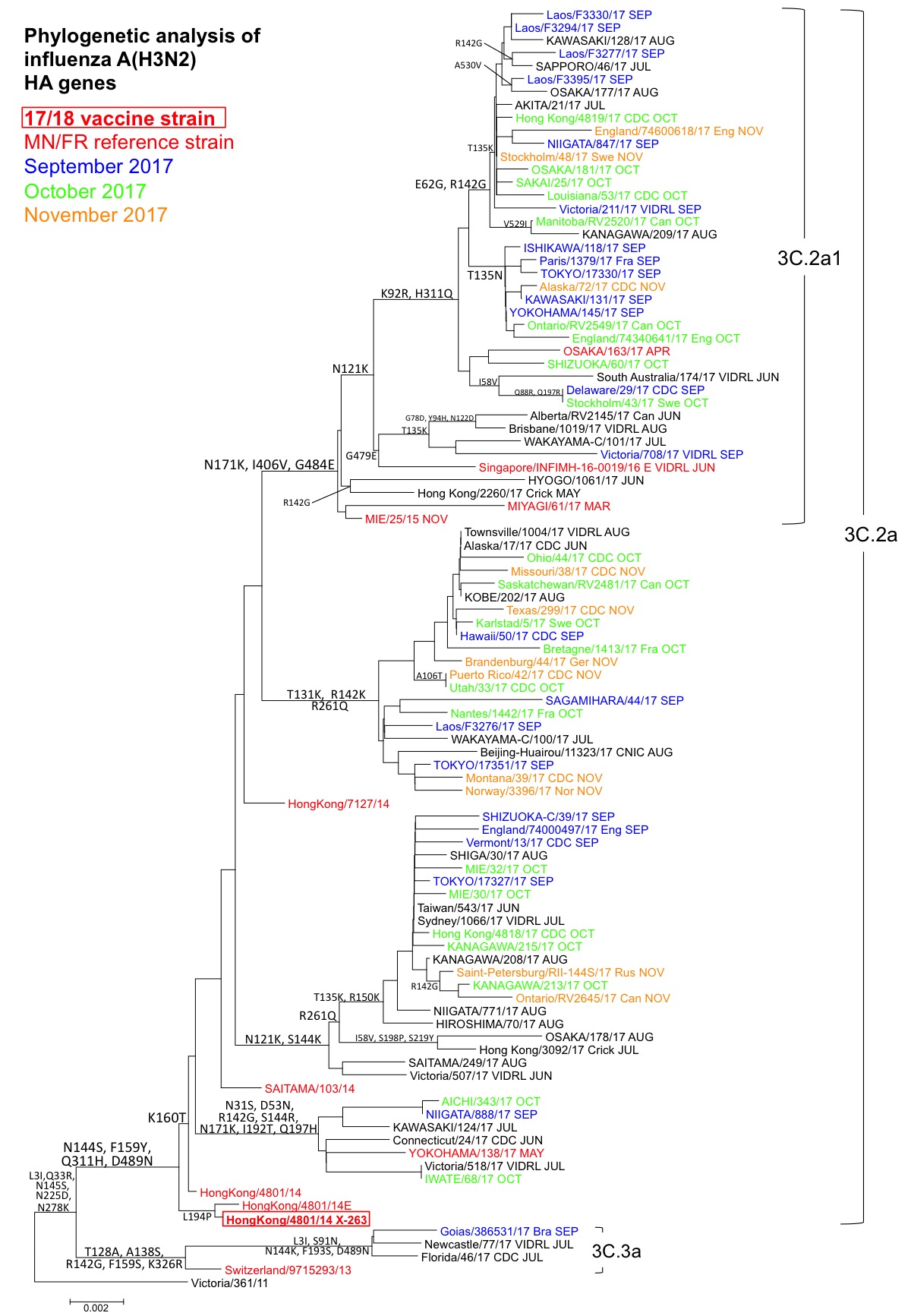
A(H3N2):最近の流行株はHA遺伝子系統樹上でサブクレード3C.2あるいは3C.3に属している。3C.2内にはさらに、2016/17シーズンのワクチン株A/Hong Kong/4801/2014株に代表されるサブクレード3C.2a(L3I, N144S, F159Y, K160T, Q311H, D489N)が形成されており、解析した流行株は全てこの3C.2aに属した(図2)。また3C.2a内にはサブクレード3C.2a1(N171K, I406V, G484E)が形成されている。3C.2a1を除く3C.2a内には、大きくN121K, S144K群、T131K, R142K, R261Q群、N31S, D53N, R142G, S144R, N171K, I192T, Q197H群が形成された。3C.2a1ではN121K群が大きな割合を占め、さらにこの群の中でG479E群およびK92R, H311Q群に分かれた。K92R, H311Q群にはE62G, R142G群およびT135N群が形成された。また3C.2a1内には他にも集団が形成されており、最近のA(H3N2)のHAは遺伝子的に多様化が進んでいる。
|
{kind=link}
|
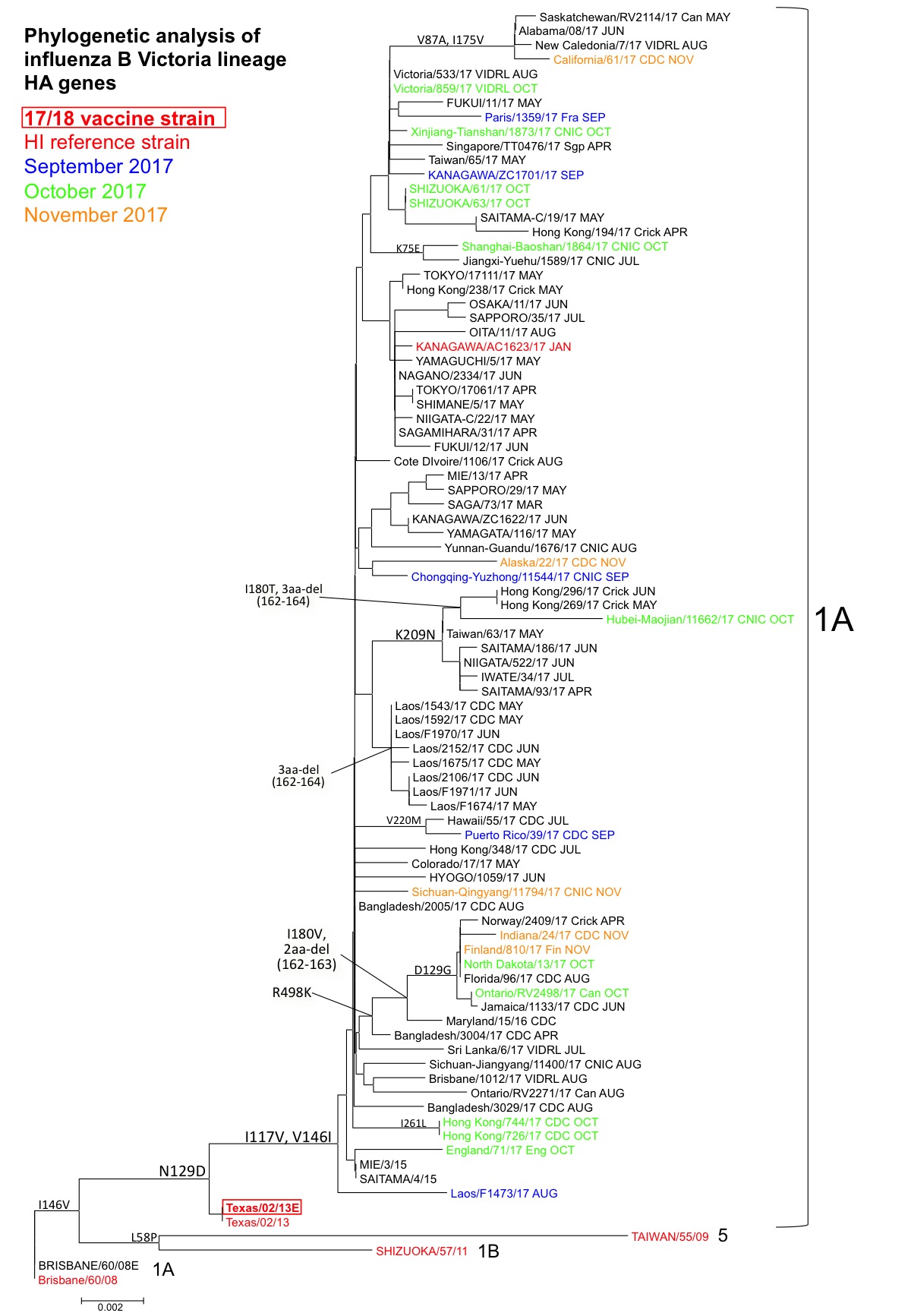
B (ビクトリア系統):解析した株は全て、HA遺伝子系統樹上のクレード1A(共通アミノ酸置換N75L、N165K, S172P、代表株:B/Brisbane/60/2008株、B/Texas/02/2013株)内の、共通アミノ酸置換N129D, I117V, V146Iを有する集団に属していた(図3)。その中で複数の群が形成されており、遺伝子的には多様化が進んでいる。なお現在までに、北米を中心に一部の海外で報告されている抗原性変異株[共通アミノ酸置換R498K, I180V,および2アミノ酸欠損(162, 163)]、香港で検出された抗原性変異株[共通アミノ酸置換K209N, I180Tおよび3アミノ酸欠損(162, 163, 164)]、およびラオスで検出された抗原性変異株[3アミノ酸欠損(162, 163, 164)]は、日本では検出されていない。
|
{kind=link}
|
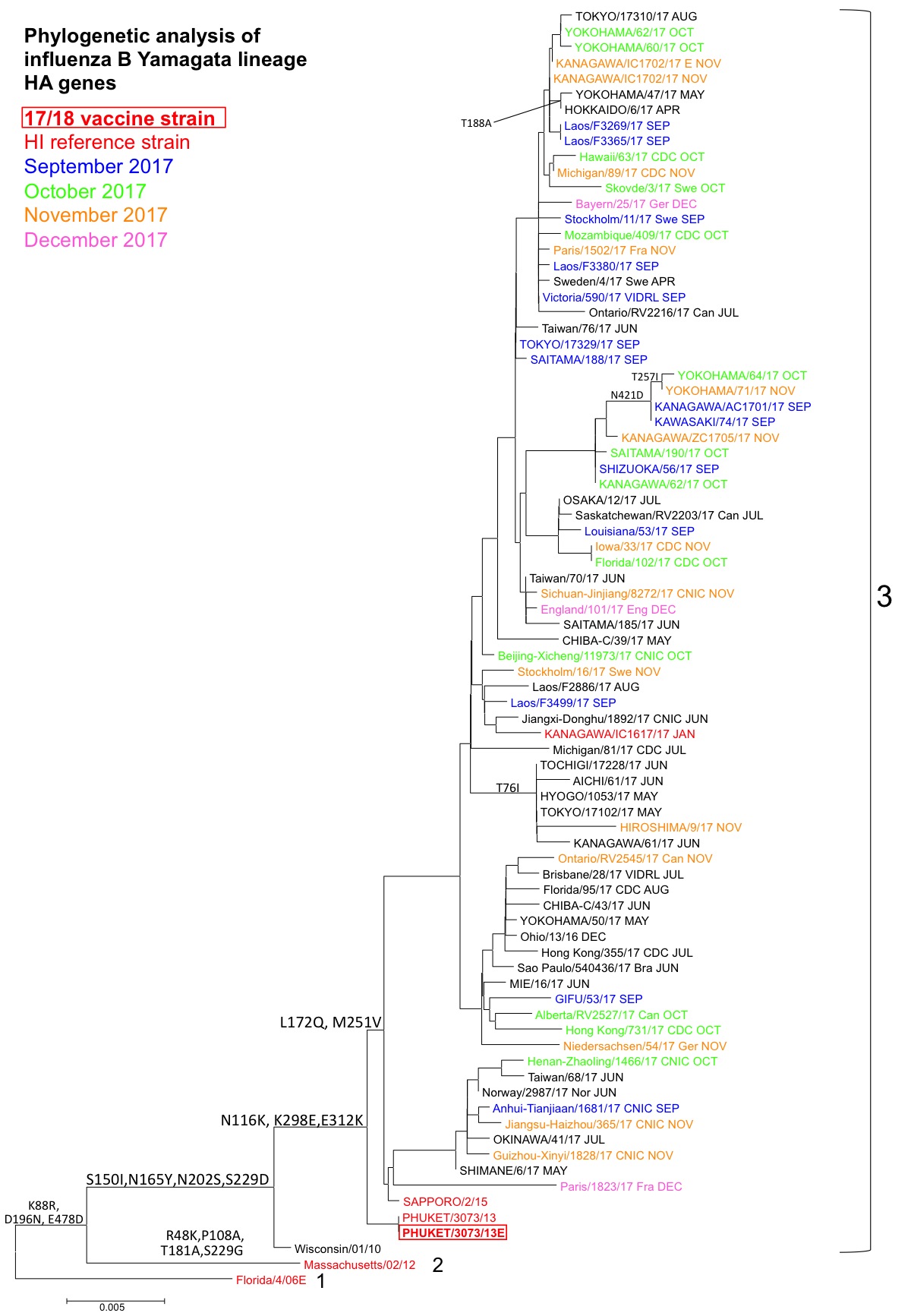
B (山形系統):解析株は全て、HA遺伝子系統樹上のクレード3(共通アミノ酸置換S150I, N165Y, N202S, S229D)内でB/Phuket/3073/2013株を代表とする群(共通アミノ酸置換N116K, K298E, E312K)内のL172Q, M251V群に属した(図4)。また、共通アミノ酸を持たないが複数の集団を形成しており、遺伝的には多様化が進んでいる。
|
{kind=link}