バンコマイシン耐性腸球菌(vancomycin-resistant enterococci: VRE)の薬剤耐性機構について
(IASR Vol. 42 p157-158: 2021年8月号)
細菌の細胞壁合成過程とバンコマイシンの作用機構
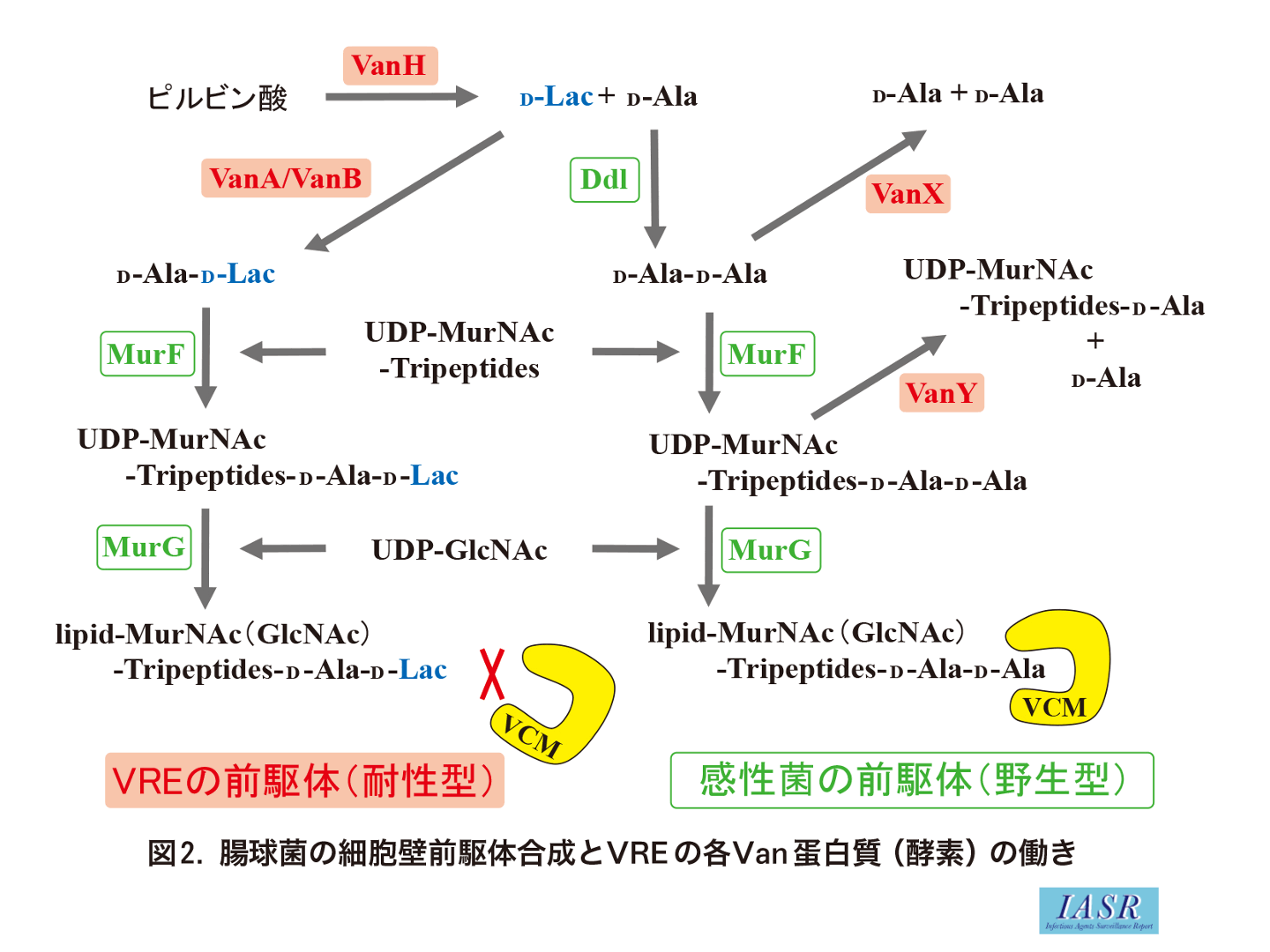
バンコマイシンを含め, グリコペプチド系薬は細菌の細胞壁の合成を阻害する抗菌薬である。細菌の細胞壁物質はペプチドグリカンで, 2種類の糖, N-アセチルムラミン酸(MurNAc)とN-アセチルグルコサミン(GlcNAc)の繰り返し結合による直列の鎖が, ペプチドで架橋されている構造をとっている。すなわち細胞壁は糖鎖を縦糸とすると, ペプチドによる架橋を横糸とする網目構造をしている。この細胞壁が合成される時, 最初に細胞質内で細胞壁前駆体(ムレインモノマー)が作られる。まず糖鎖の構成成分の基となるUDP-MurNAcにアミノ酸が順次結合し, 最終的に5個のアミノ酸によるペプチド鎖(ペンタペプチド)が付加され, さらにもう1つの糖であるGlcNAcが結合する。その結果, 最終的にムレインモノマーであるlipid-MurNAc(GlcNAc)-L-Ala1-γ-D-Glu2-L-Lys3-D-Ala4-D-Ala5が細胞膜の内側で形成される。なお, ペンタペプチドのD-Ala4-D-Ala5部分は, リガーゼ酵素Ddl(ddl遺伝子産物)の働きによって2分子のD-AlaからD-Ala-D-Alaが先に合成され, これがペプチド鎖末端の-L-Lys3に付加することでペンタペプチドが形成される。このムレインモノマーが細胞膜外へ運ばれ(細胞膜上で反転), 細胞壁架橋酵素であるペニシリン結合蛋白PBPの働きによって, 合成中のペプチドグリカンと前駆体との糖鎖間およびペプチド鎖間での結合がそれぞれ起こり, 細胞壁が完成する。このペプチド鎖同士が結合する時にペンタペプチド末端の5番目のD-Ala5が切られ, 4番目のD-Ala4が他のペプチド鎖と結合することにより架橋される。
グリコペプチド系薬はグラム陽性菌に有効で, 菌の細胞膜外において細胞壁前駆体ムレインモノマーのペンタペプチド末端のD-Ala4-D-Ala5部分に特異的に結合する。そのためPBPによるペプチド結合(架橋反応)が阻害され, 細胞壁合成が停止する。グラム陰性菌においては薬剤が外膜を通過することができず, その作用点であるペプチドグリカン層に到達できない。そのためグラム陰性菌は, バンコマイシンに対して自然耐性である。
VREのバンコマイシン耐性機序と耐性遺伝子
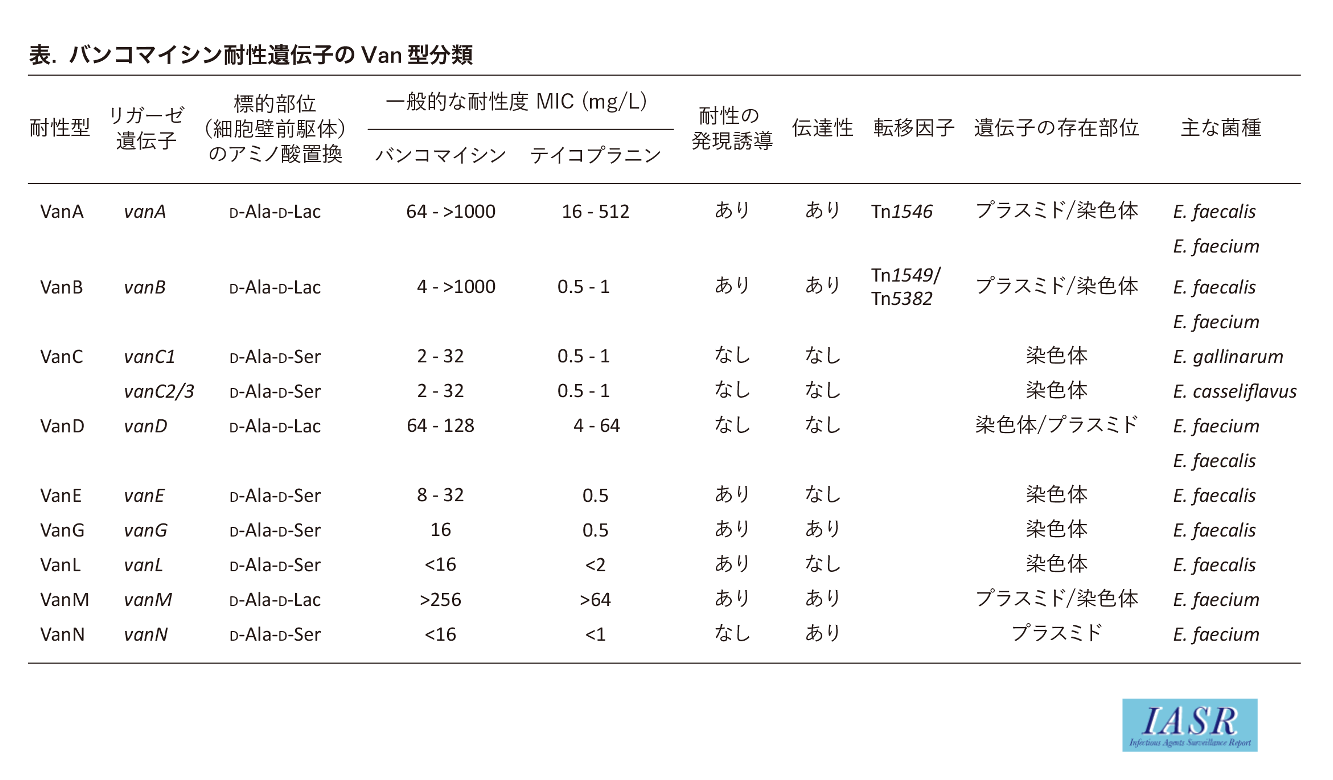
1)VREでは細胞壁前駆体の-D-Ala4-D-Ala5部分の5番目のD-Ala5がD-lactate(乳酸, 図表ではLacと表記), またはD-Ser(セリン)に置換され, -D-Ala4-D-lactate5あるいは-D-Ala4-D-Ser5となっている(表)。グリコペプチド系抗菌薬は, これらの前駆体に結合できないために菌の細胞壁合成が阻害されず薬剤耐性となる。D-Ala-D-lactateを形成する耐性型(後述)の方がD-Ala-D-Serを形成する他の耐性型よりも薬剤親和性が低いために, 高度耐性(高MIC値)となる。ペプチド鎖による架橋形成時には末端の-D-lactate5あるいは-D-Ser5は切り離されるので, でき上がったVREの細胞壁は通常の細胞壁と変わりがない。VREとして複数の耐性型(Van型)が報告されているが, 高度耐性を示す耐性型(-D-Ala4-D-lactate5構造の前駆体合成)としてVanA型, VanB型, VanD型, VanM型が知られており, それらの起源はグリコペプチド系抗菌薬を産生する微生物(放線菌類)と考えられている(表, 図1)。VREの高度バンコマイシン耐性は, 可動因子(伝達性のプラスミドやトランスポゾン)上に存在するこれら特異的な耐性遺伝子を外来性に獲得したことによるもので, 単に抗菌薬投与や突然変異によって高度耐性VREが生じることはない。実際の臨床においてはVanA型およびVanB型が多く分離され, また菌種としてはEnterococcus faeciumが主で, 次いでE. faecalisが多く分離される。他のVan型は-D-Ala4-D-Ser5構造の前駆体を合成するVREであり, 高度耐性を示さないことから臨床上で問題となることは少ない。
{kind=link}
{kind=link}
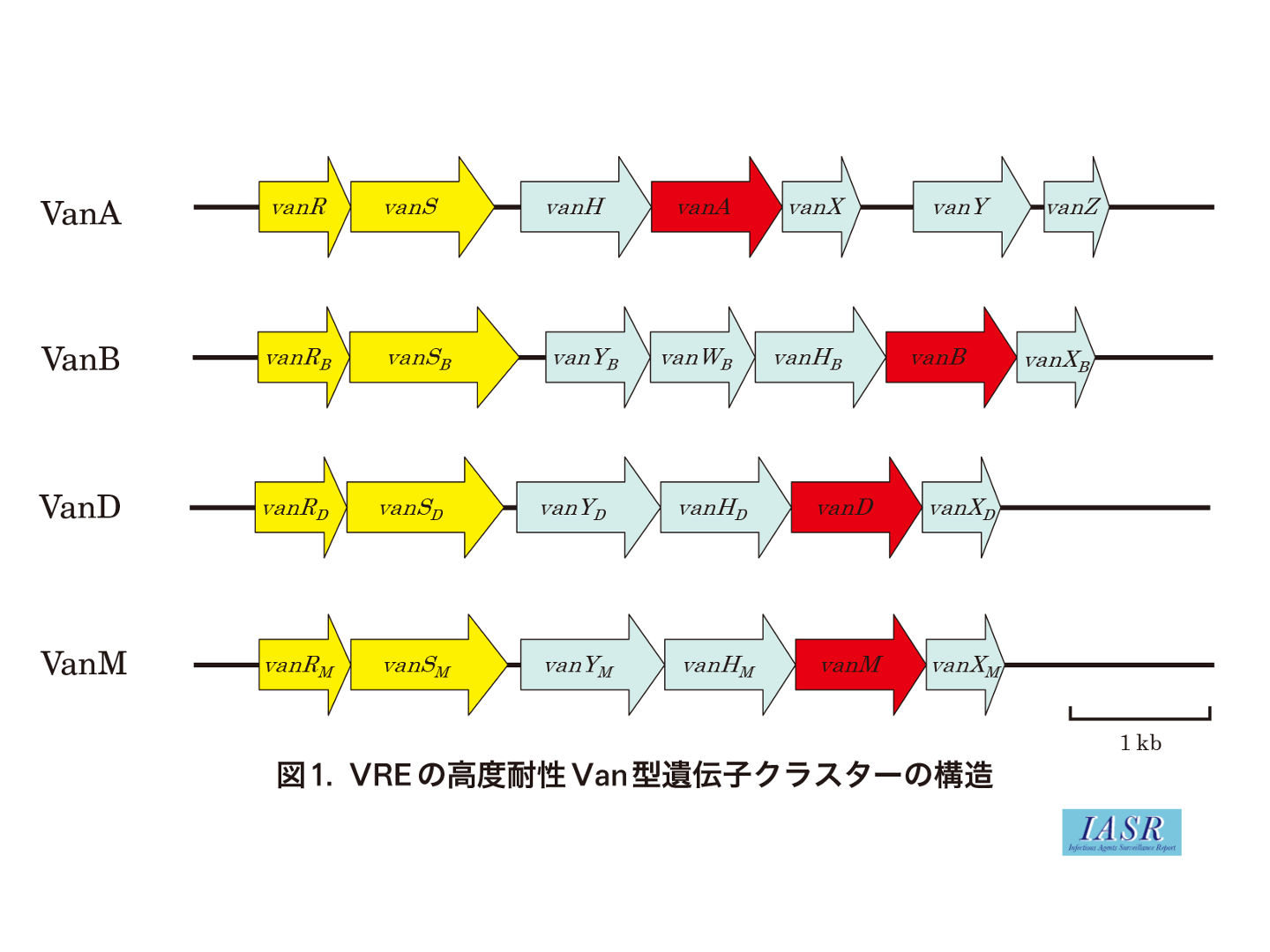
各Van型VREの基本となる保存された耐性遺伝子は, vanRSHAXYZである(図1)。vanRとvanSはvanHAX発現のための調節遺伝子, vanHAXYはバンコマイシン耐性のための遺伝子である(図2)。VanHは酸化還元酵素で, NADP(H)を酸化し, ピルビン酸を還元しD-lactate(乳酸)を生産する。VanAはD-AlaとD-lactateの結合酵素(リガーゼ)で, これによりD-Ala-D-lactate が形成される。このdipeptideがUDP-tripeptideに結合され, UDP-tripeptide-D-Ala4-D-lactate5(またはUDP-tripeptide-D-Ala4-D-Ser5)ができる。VanXは正常なdipeptideであるD-Ala-D-Alaを分解し, バンコマイシン感性となる(野生型)前駆体の産生を抑える。VanYは正常な前駆体であるUDP-tripeptide-D-Ala4-D-Ala5から末端のD-Ala5を切り離し, VanXと同様に野生型前駆体の産生を抑える。VanY, VanXによって切り出されたD-Alaは-D-Ala4-D-lactate5合成のための基質として再利用される。VREはこれらVan蛋白質の協調的な働きによって, バンコマイシンを含むグリコペプチド系抗菌薬の存在下でのみ耐性型の細胞壁前駆体を合成して薬剤耐性となる。一方で, 抗菌薬の存在下においても染色体性のddl遺伝子(Ddlリガーゼ酵素)は発現するために, ペプチド末端が野生型(バンコマイシン結合可能)-D-Ala4-D-Ala5の細胞壁前駆体も合成されている(図2)。実際のVREにおいてはこの野生型と耐性型の前駆体の量比も耐性度に影響すると考えられている。
{kind=link}
参考文献
- 富田ら, 薬剤耐性菌制御のための教育セミナー:資料集:108-121
http://yakutai.dept.med.gunma-u.ac.jp/project/5th_KyouikuSeminar(40.5MB)v2.pdf