わが国のE型肝炎分子疫学情報(2016年~2021年第42週)
(IASR Vol. 42 p277-279: 2021年12月号)
E型肝炎は潜伏期間が長いことから, 聞き取りによる感染源の同定は困難であり, 感染源の共通性の検討には, 患者の糞便等から分離されるウイルス株についての分子疫学的手法による解析が有用である。E型肝炎の流行状況を調査するため, 厚生労働省は「E型肝炎発生時の検体の確保等について」〔2016(平成28)年8月16日, 健感発0816第3号, 生食監発0816第2号〕を発出し, 各自治体宛にE型肝炎の発生届を受理した際に, 患者検体の確保, もしくはウイルス解析情報の国立感染症研究所(感染研)への提出を依頼している。提出された患者検体は病原体検出マニュアルに従い, ウイルスRNAの抽出とE型肝炎ウイルス(hepatitis E virus: HEV)のORF2内の遺伝子配列を決定している。感染研で決定した遺伝子配列, および自治体から提供された遺伝子配列により系統樹を作成し, 感染症発生動向調査(NESID)と組み合わせた分子疫学的な積極的疫学調査を行っている。
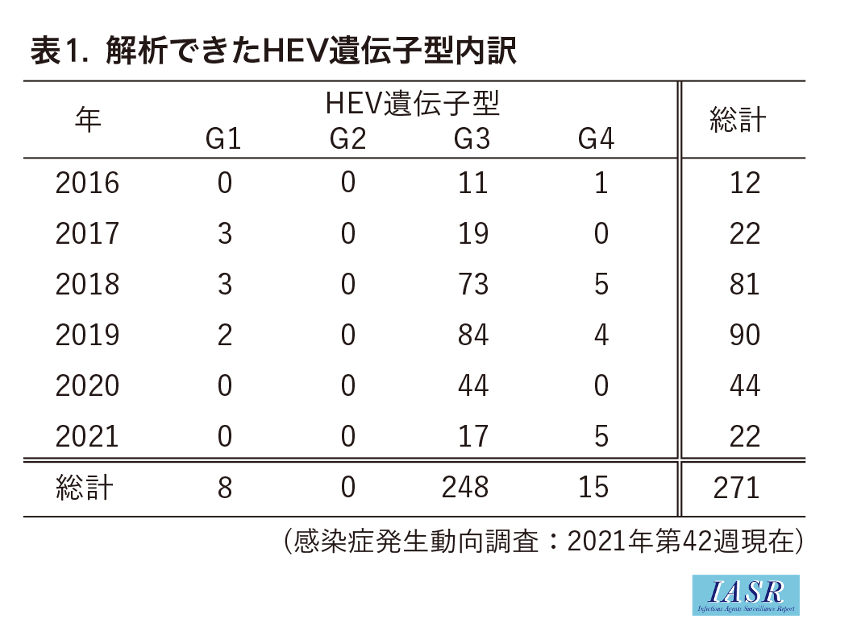
積極的疫学調査のための患者検体もしくはウイルス解析情報の収集は2016年より行われ, これまでにNESIDに紐付けられた検体が370例集まり, そのうち271例で塩基配列情報ならびに遺伝子型が決定された。
NESIDによると, 2016年~2021年第42週までの年間のE型肝炎届出数は, 300例を超えており, 近年は年間450-500例にのぼる。性別年齢分布の比較では, 男女比は3:1と男性での届出が多い。男女ともに年齢分布は10代後半~100歳代まで幅広い年齢で届出されるものの, 40~80代が大半を占め, 各年の年齢中央値は60歳前後である。この傾向は2016年以降各年で変化は認められない。またE型肝炎の臨床症状としては, 黄疸や肝腫大, 肝機能異常, 食欲不振, 全身倦怠感, 発熱等が挙げられ, 無症候や自覚症状がない場合もあるが, これらの発現割合についても各年で特段の変化は認められない。
NESIDの調査項目において各年で大きな差は認められないことから, ここでは2016年以降, NESIDと紐付けることができた塩基配列情報271例についてまとめることとした。
HEVの遺伝子型(genotype)については, E型肝炎患者からは主に4つのgenotype(G1-G4)が検出され, それぞれのgenotypeは地理的分布が異なっている。G1はアジア・アフリカ諸国のE型肝炎流行地域, G2はメキシコおよびナイジェリア, ナミビア, エジプトなどのアフリカ諸国, G3は世界中に広く分布し, G4は中国, 台湾, ベトナム, インド, インドネシア等のアジア地域を主とし, 日本にも分布している。G1, G2は従来の流行性肝炎の原因となっているのに対して, G3, G4はブタやイノシシ等の動物が主要なリザーバーと考えられ, ヒトでの散発的なHEV感染の原因となっている。わが国においてはG3の検出が多いが, 北海道ではG4が多く検出されることが報告されている。
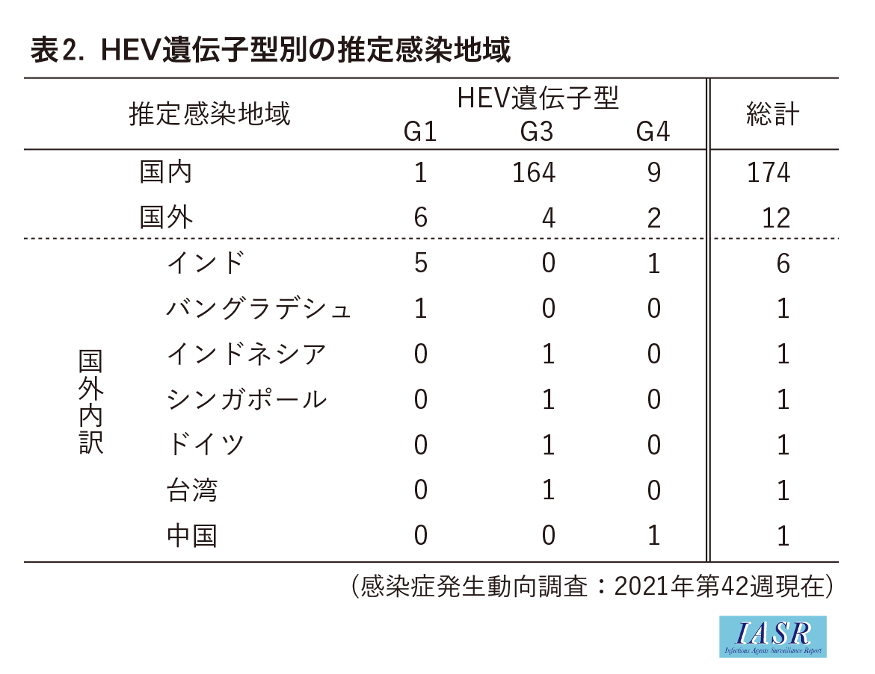
2016年以降の積極的疫学調査で明らかになった271例のgenotype内訳はG1が8例, G2が0例, G3が248例, G4が15例であった(表1)。推定される感染地域として国内174例, 国外12例の記載があり, genotype別で比較すると, G1は国内1例, 国外6例, G3は国内164例, 国外4例, G4は国内9例, 国外2例であった。G3とG4は国内での感染例が多かったのに対して, G1は国外での感染例の方が多く, 感染地域はG1の侵淫が報告されている東アジア地域のインドおよびバングラデシュであった(表2)。
{kind=link}
{kind=link}
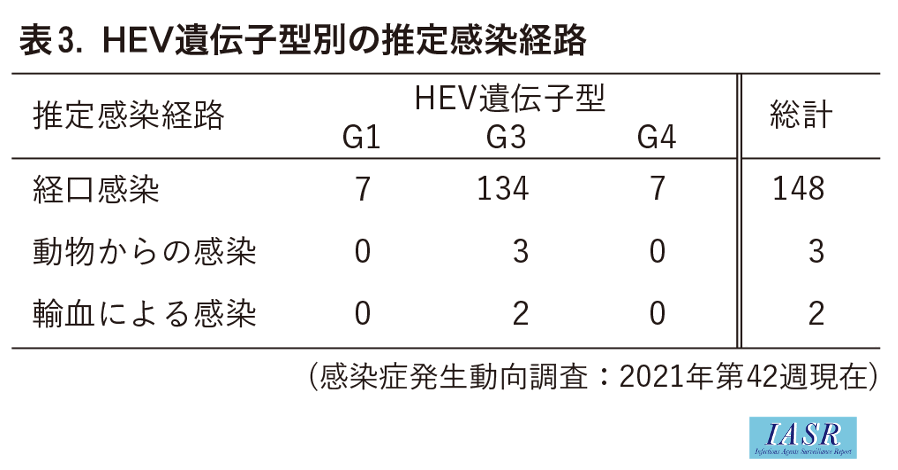
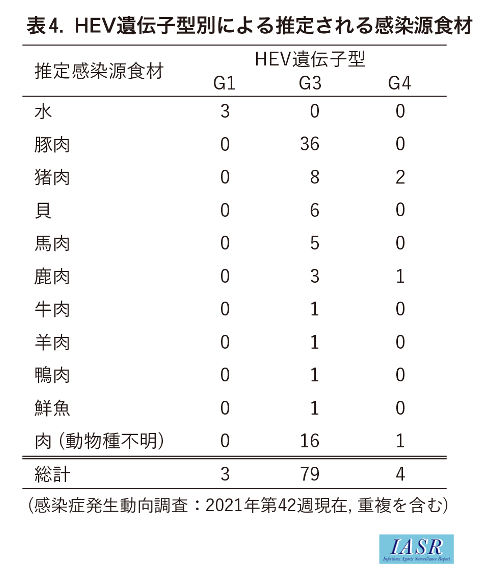
Genotype別に推定感染経路を比較すると, G1は経口感染のみ7例の記載があり, そのうち3例で詳細の記載があり, すべて水によるものであった。G3は経口感染134例で, そのうち72例について詳細の記載があり, ブタ(肉やレバーを含む)の喫食が36例, イノシシ8例, 貝6例, ウマ5例, シカ3例, ウシ, ヒツジ, カモ, 鮮魚が各1例ずつ, 動物種不明の肉類 (生肉, 焼肉など)16例であった (重複を含む)。それ以外に, 野生動物や飼育, 調理による感染3例, 輸血による感染2例が記載されていた。G4は経口感染7例の記載があり, そのうち4例で詳細の記載があり, イノシシ2例, シカ1例, 動物種不明の肉類(焼肉)1例であった(表3,4)。G1はこれまでブタやイノシシへの感染は報告されておらず, 今回の調査でも水による感染が疑われた。一方でG3, G4はブタやイノシシ等への感染が報告されており, 今回の調査でも豚肉や猪肉の喫食による報告例が多数を占め, あらためて人獣共通感染症としてのリスクを浮き彫りにした。ただしNESIDに記載された感染地域, 感染経路は推定であり, 特定されたものではないことに留意する必要がある。
{kind=link}
{kind=link}
次に系統樹によるクラスター解析を行ったところ, 積極的疫学調査が開始された当初は同一クラスターに分類される株は少なく, 散発例がほとんどであったが, 報告例が蓄積されたことで, 近年は同一クラスターに分類される株もみられるようになってきた。同一クラスターに分類された株についてNESIDと照らし合わせたが, 推定感染年月日や推定感染地域, 推定感染経路はまちまちであり, 共通点はなかなか見出せていないのが現状である。
患者検体もしくはウイルス解析情報の収集は, 2016年の開始以降徐々に増加してきている。塩基配列データが蓄積されるほど解析の精度も向上すると考えられるが, 新型コロナウイルス感染症(COVID-19)による影響により2020, 2021年の塩基配列データの集積数は著しく減少している。今後はより幅広く都道府県, 自治体の協力を得ることにより, 本積極的疫学調査を充実させていくことが重要であると考えている。
謝辞:貴重な検体や塩基配列情報をご提供いただきました地方衛生研究所ならびに保健所の方々に心より感謝申し上げます。