お知らせ
感染症情報
研究・検査・病原体管理
サーベイランス
刊行・マニュアル・基準
- 詳細
国立感染症研究所 免疫部 主任研究官 大西 和夫
平成21年6月22日
免疫部
大西和夫(内線2129)
「免疫機能に関する研究:抗体産生B細胞の発生・分化機構に関する研究」
感染症における抗体の役割りについて
細菌やウイルスなどの病原体が体に侵入してきた時におこる生体防御反応には、大きく分けて「自然免疫」と「獲得免疫」があります。「自然免疫」は、幅広い病原体に対して生体がもともと持つ抵抗性で、昆虫や植物も含めた生物界全般に備わった免疫機構を指します。これに対して「獲得免疫」は、例えばインフルエンザ・ウイルスやジフテリア毒素などが体に侵入した時に、それらの病原体に固有の「免疫的な記憶」を獲得することにより生体を防御する機構で、進化的には脊椎動物にのみ発達しています。「獲得免疫」機構の中心的な役割を演ずる分子の一つが「抗体」です。
抗体は、体に侵入してきた病原体などの異物(これを「抗原」という)を厳密に見分けることができます。これを抗体の「抗原認識特異性」といいます。たとえば、インフルエンザ・ウイルスが感染して体内で増殖した時、このウイルスの表面にある分子の様々な細かい構造を認識して結合できる様々な抗体が何百種類も出来てきます。このうち、いくつかの抗体はウイルスが細胞に取り付いて侵入するのを阻害することが出来ることから、生体からのウイルス排除に役立ちます。このような抗体を中和抗体と呼びます。後で述べますが、抗体分子をつくる細胞はB細胞と呼ばれるリンパ球で、一部のB細胞は非常に長寿命で、次に同じ病原体が侵入してきた時に速やかに有効な抗体を産生することが出来ます。これを「B細胞免疫記憶」と呼びます。ここでは詳しく述べませんが、他に「T細胞免疫記憶」と呼ばれる現象があり、両者を合わせて「免疫記憶」と呼びます(最近の研究ではNK細胞にも免疫記憶があるとする報告があります)。
感染症の予防に中心的な役割を果たす「ワクチン」は、不活化(弱毒化)した病原体を接種することにより、その病原体に対する「免疫記憶」を植え付けるものです。このことから、ワクチン等の外来抗原により、生体の免疫システムにおいてどのように免疫記憶が獲得され、維持されるかを知ることは、より機能的なワクチンを開発する上での基盤技術になります。また、抗体の厳密な「抗原認識特異性」は、感染病原体の同定に使われる「診断薬」の作製に応用されています。例えば、インフルエンザのような初期症状を呈した時、それがどのようなタイプのインフルエンザ・ウイルスによるものなのか、もしくは近年流行したSARSウイルスによるものなのかを診断する体外診断薬キットの作製に使われます。さらに、最近特に注目されている「抗体医薬」は、抗体の抗原認識特異性を治療薬に応用したもので、癌や免疫関連疾患の標的分子をピンポイントで制御出来る点で非常に優れており大きな期待が集まっています。
私たちは、体の中で抗体がどのようにつくられ、どのように働き、またどのように免疫記憶として免疫システムに固定されていくかを研究することにより、ワクチン・診断薬・治療薬の開発に役立つ基盤技術への応用を目指しています。
以下に、私たちが行っている基礎研究の一つを紹介します。
抗体の抗原認識多様性を管理するプレB細胞レセプター
上に述べたように、抗体(免疫グロブリン、Ig)は、生体防御の第一線に働く重要な分子であり、その機能は様々な外来異物の認識を可能にする抗原認識部位(レパトア)の多様性に依存しています。この多様性が低ければ、認識・排除できない外来抗原のために個体の生存が脅かされることは容易に想像でき、実験的にも証明されています。免疫グロブリンの抗原認識部位の多様性は、利根川進博士によって示されたように遺伝子組み換えによるV、D、J領域の再構成と、その時に起こるN配列の挿入によって生み出されます。このランダムな遺伝子断片の組み合わせと挿入変異によって計算上は無限に近い多様性が生み出されますが、その多様性はプレB細胞レセプター(preBCR)の選択を受けて免疫システムに組み込まれることが最近明らかになりつつあます(図1)。
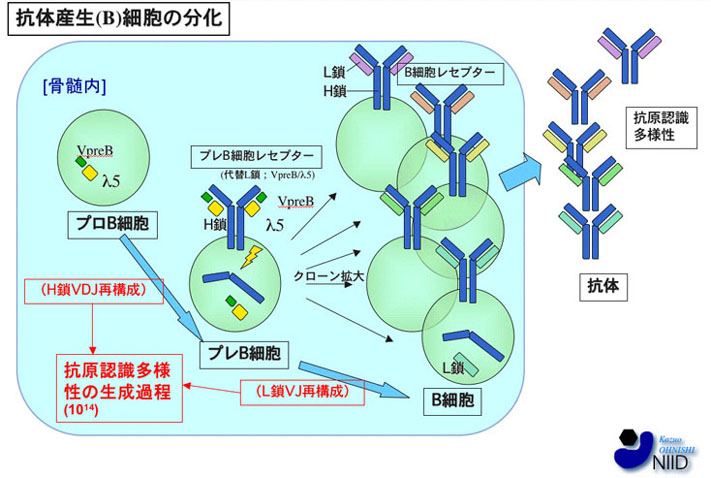
図1:B細胞分化の略図
骨髄においてプロB細胞期に抗体H鎖遺伝子座のVDJ再構成が開始され、プレB細胞期にはH鎖のみが発現している。この時期にVpreBとλ5から成る代替L鎖(Surrogate Light-Chain)が結合してプレB細胞レセプター(PreBCR)が形成されて細胞表面に発現する。PreBCRからのシグナルは細胞分裂を誘導しクローンの拡大を引き起こす。一つの抗体H鎖のclonotypeを持って増殖した細胞群は各々独立にL鎖遺伝子再構成を行うので、H鎖/L鎖との組み合わせの数を増やすことができる。この後、B細胞レセプター(BCR)が細胞表面に発現するとともに骨髄を出て多様な抗原認識多様性を持った抗体群が形成される。
プレB細胞レセプターは、代替L鎖と呼ばれる2つの分子(λ5とVpreB)が抗体L鎖様の構造をとり、遺伝子再構成の結果新しく発現してくる抗体H鎖と結合することによってpreBCRとしてB細胞表面に発現します。この代替L鎖の役割りについては以下のように考えられています。B細胞の初期分化の過程で抗体H鎖遺伝子再構成がまだ起っていない時期、すなわちプロB細胞期においては、VpreB、λ5およびIg-α、Ig-βといったpreBCRを構成する分子はすでに発現されており、遺伝子再構成の結果生まれる新生H鎖の発現を待っているかのような状況になっています。図2にあるように、抗体H鎖遺伝子再構成の際、V→DJ組み換えは、コドンの読み枠の関係から成功率が約3分の1であるが、運良くうまく行った場合にはもう一方の染色体上にあるIg遺伝子座のV→DJ再構成はキャンセルされます(対立遺伝子排除、allelic exclusion)。
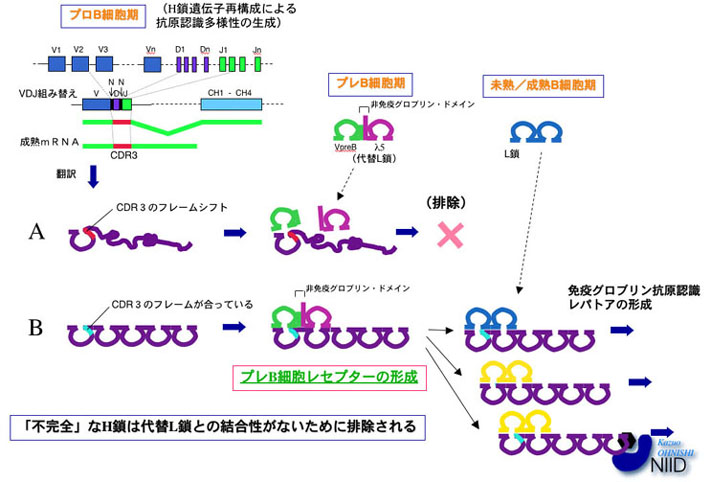
図2:抗体H鎖抗原認識多様性の生成と代替L鎖による新生H鎖の品質管理
抗体H鎖抗原認識部位のうち最も重要なCDR3の多様性はH鎖V→DJ再構成とN領域の挿入により形成される。この時、コドンの読み枠がずれる場合には、CH1以下が異常なタンパク質となり、代替L鎖が結合出来ない事からPreBCRが形成されず、細胞は分化の次のステップに進むことが出来ない(A)。読み枠が合っている場合には、代替L鎖が結合してPreBCRを形成し、そのシグナルによりクローンの拡大を引き起こす(B)。この意味で代替L鎖は新生H鎖の品質管理をしているといえる。PreBCRの活性化には、代替L鎖上のnon-Igドメインが深く関わっている。
これにより、一つのB細胞が一つの抗原認識特異性を持つことが保障されます。逆に、最初のV→DJ再構成がうまく行かなかった場合は、対立遺伝子座にそれが誘導され、もう一度完全なμH鎖を作る機会が与えられます。2度のチャンスに失敗したB細胞はアポプトーシスを起こし排除されると考えられています。代替L鎖はこの過程において新生H鎖の品質管理を行っていると考えられています(図2)。すなわち、先に述べたように、プロB細胞後期に起こるH鎖遺伝子再構成は、コドンの読み枠の関係から成功率は約3分の1であり、3分の2はVHの下流に異常な蛋白質(Igドメイン構造をとらないもの)が出来てきます。VpreBとλ5からなる代替L鎖は、各々の持つIgドメインでH鎖のVHドメインとCH1ドメインを認識することにより新しく出来てきたH鎖のドメイン構造をチェックします。H鎖が正常なIgドメイン構造を持つならば代替L鎖は通常のL鎖と同様な様式でH鎖と結合し、preBCRを形成します。もし、V→DJ再構成がうまく行かず翻訳された蛋白質が異常な構造をとる場合には、Igドメイン間の相互作用を持てないために代替L鎖は結合できず、preBCRは形成されません。前者の場合、形成されたpreBCRは膜表面に発現して、対立遺伝子排除、クローンの拡大、L鎖遺伝子座再構成の誘導等、再構成に成功したH鎖を最大限に免疫システムに組み込もうとする合目的的な現象を引き起こすシグナルを送ると考えられています。preBCRを形成できない幾つかのノックアウトマウスではB細胞の分化がプレB細胞期でブロックされており、その後の分化の進行が強く阻害されます。
プレB細胞レセプターの自動的活性化機構
では、「どのようにこのB細胞の初期分化で中心的な役割を果たすpreBCRが活性化されるか」という問題に関して、最近の私たちの研究で、代替L鎖の特定のドメインがあたかも抗原のような役割をしてレセプターの自己架橋・自己活性化を誘導するという興味深い知見を得ています。λ5分子は、C末側の3分の2は抗体lL鎖のJ領域+定常領域に非常に高いホモロジーを持ちます。N末側3分の1は、既知の蛋白とのホモロジーがなく、非免疫グロブリン(非Ig)領域と呼ばれます。一方のVpreBは、N末側の4分の3はL鎖可変部と高いホモロジーを持ちますがC末側4分の1はホモロジーがなく、やはり非Ig領域と呼ばれます。VpreBとλ5の非Ig領域は、興味あることにL鎖の抗原認識部位であるCDR3に相当する部位に非常に大きな構造を造ることになります(図3)。
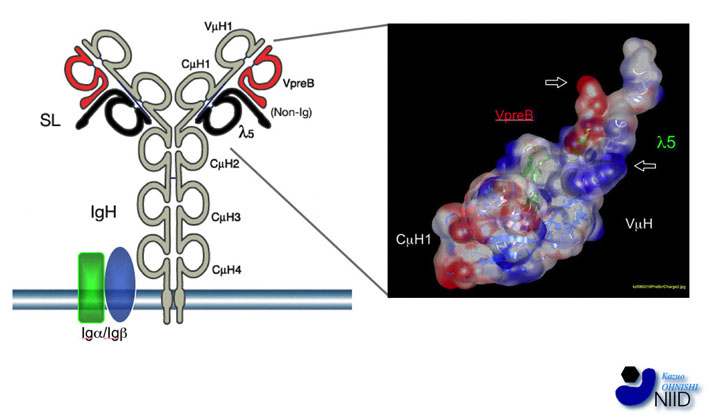
図3:プレB細胞レセプターの構造と非Ig領域
(左図)プレB細胞レセプター(preBCR)には、L鎖の代わりに代替L鎖(λ5とVpreB分子からなる)が結合している。λ5分子は、C末側の3分の2はL鎖にホモロジーが高く、N末側3分の1はホモロジーがないため非Ig領域(Non-Ig)と呼ばれる。VpreBは、N末側の4分の3はL鎖可変部と高いホモロジーを持つがC末側4分の1はホモロジーがなく、やはり非Ig領域と呼ばれる。(右図)VpreB/λ5とVμH・CμH1鎖が形成する構造をホモロジーモデリング法により予測した。VpreBとλ5の非Ig領域は、CDR3に相当する部位に大きな構造を造る(矢印)。分子動力学的手法を用いて非Ig領域構造の分子運動のシミュレーションを行った結果、この領域は相当フレキシブルな自由度の高い構造であることが示唆されている。非Ig領域には、電荷を持つアミノ酸が集中しており、その電荷に基づく静電的な相互作用によって隣り合ったPre-B細胞レセプター同士が自動的に架橋するメカニズムが示唆される。
λ5とVpreBの非Ig領域には、電荷を持つアミノ酸が集中しており、その位置も進化的に保存されています。そこで、突然変異の導入によりこれらの電荷を持つアミノ酸を打ち消したpreBCRを作製してその機能を調べたところ、レセプターの活性化がほとんど起こらなくなることを見つけました。このことは、λ5の非Ig領域に集中した荷電アミノ酸がこのレセプターの活性化モチーフであることを強く示唆します。つまり、生理的な条件下ではpreBCRの架橋と活性化は細胞外のリガンドを必要とせずに自発的に起こる事を示唆しています(図4)。

図4:プレB細胞レセプター活性化機構のモデル
λ5とVpreBのnon-Igドメインは、免疫グロブリンのCDR3に相当する部分に大きな構造を作り、λ5のnon-Igドメインには7個のアルギニンが進化的に保存され、+の電荷を付与している。VpreBのnon-Igドメインにある電荷との相互作用は不明だが、それによりpreBCRの架橋と活性化は自発的に起こり、外来のリガンドを特に必要としないことが強く示唆される。preBCRの機能を考えると、それが細胞外の環境に向かったセンサーではなく、細胞内に向かったセンサー、つまり、新しく発現したm鎖タンパク質の品質管理であることを考えると、このレセプターの自己架橋と自己活性化は非常に合目的な機構であると考えられる。
preBCRの機能が新生H鎖の品質管理であることを考えると、それがレセプターの本来の機能である細胞外の環境に向かったセンサーではなく、細胞内に向かったセンサーであることが判ります。また、このレセプターが外来のリガンドを必要とせずに自己架橋・自己活性化する機構は、preBCRシグナルの発生がH鎖のclonotypeに非依存的である事を保証します。このことは、H鎖の抗原認識多様性を保存して免疫系に組み込む上で非常に合目的的な機構であると考えられます。
本研究の展望と応用
上述した様に、preBCRの形成は新生H鎖を免疫系に組み込む上で巧妙な戦略を実現しているが、その基盤は新生H鎖と代替L鎖の結合性に依存している事が理解できる。では、どのような新生H鎖でも代替L鎖と結合できるのだろうか?H鎖のCDR3の構造によっては、L鎖や代替L鎖の結合無しに細胞表面に発現するL鎖非依存性H鎖の存在が報告されている。さらに重要なのは、H鎖V→DJ再構成に成功した、つまり正常なIgドメイン構造を持つにも関わらず代替L鎖と結合できないH鎖が予想外に高頻度に存在している証拠が報告されて来ていることである。この事は、代替L鎖とH鎖の結合様式が結果として形成される抗体の抗原認識レパトアを著しく規定(または拘束)し、免疫系の抗原認識多様性の形成に深く関与していることを示唆している。この拘束の分子構造的基盤を明らかにし、代替L鎖とH鎖の結合性をコントロールできれば、抗体の抗原認識特異性を飛躍的に高める全く新しい技術の開発につながると考えている。
これまで、本研究を通して得た知識・技術をもとに、モノクローナル抗体を用いた重症急性呼吸器症候群(SARS)ウイルスの迅速診断キットの開発、牛海綿状脳症(BSE)プリオンの免疫系における増殖機構の研究、トリH5N1インフルエンザ・ウイルスの研究用モノクローナル抗体の開発などを行ってきた。今後、診断用抗体・抗体医薬・ワクチンの開発に応用できる新しい技術基盤の確立を目指して行く。
文献:
- Ohnishi, K.,‘Establishment and characterization of monoclonal antibodies against SARS-CoV' IN: SARS and Other Coronaviruses: Strategies and Protocols (ed. David Cavanagh), The Humana Press Inc., pp191-204 (2008).
- 大西和夫, 「pre-B細胞レセプターからのシグナル伝達」、臨床免疫・アレルギー科、 49,121-127 (2008).
- 大西和夫,「リンパ球分化」,免疫学ハンドブック、第2部 「免疫の分子機構」、 オーム社(2005).
- Ohnishi, K., Melchers, F., Shimizu, T.,‘Lymphocyte-expressed BILL-cadherin/cadherin-17 contributes to the development of B cells at two stages’Eur. J. Immunol.,35:957-963 (2005).
- Ohnishi, K., Sakaguchi, M., Kaji, T., Akagawa, K., Taniyama, T., Kasai, M., Tsunetsugu-Yokota, Y., Ohshima, M., Yamamoto, K., Takasuka, N., Hashimoto, S., Ato, M., Fujii, H., Takahashi, Y., Morikawa, S., Koji, I., Sata, T., Takagi, H., Itamura, S., Odagiri, T., Miyamura, T., Kurane, I., Tashiro, M., Kurata, T., Yoshikura, H. and Takemori, T., 'Immunological detection of severe acute respiratory syndrome (SARS) coronavirus by monoclonal antibodies, Jpn. J. Infect. Dis., 58:88-94 (2005).
- 大西和夫、「pre-BCRシグナルはいかにして用意されるか」、臨床免疫、41巻6号、629-637 (2004).
- Ohnishi, K., Melchers, F., ‘The nonimmunoglobulin portion of λ5 mediates cell-autonomous pre-B cell receptor signaling. Nature Immunol., 4:849-856 (2003).
- Schlissel, M., ‘How pre-B cells know when they have it right., Nature Immunol., 4:817-819 (2003).
- Ohnishi, K., Shimizu, T., Karasuyama, H., Melchers, F.,‘The identification of a nonclassical cadherin expressed during B cell development and it’s interaction with surrogate light chain.’J. Biol. Chem., 275:31134-31144 (2000).
本ホームページの内容についてのお問合せ先:国立感染症研究所免疫部
03-5285-1111(代表)より内線2131にご連絡ください