肺炎マイコプラズマの遺伝子型別法と薬剤耐性の動向
(IASR Vol. 45 p6-8: 2024年1月号)肺炎マイコプラズマ(Mycoplasma pneumoniae: M. pneumoniae)は, 種内でゲノム全体の構成と配列がよく保存されており, 分離株ごとの違いは少ない。しかし, p1遺伝子型分析, MLST(multi-locus sequence typing), MLVA(multiple-locus variable-number tandem repeat analysis), SNP(single nucleotide polymorphism)分析, ゲノム解析などの方法で分離株の型別と分類が行われている1-6)。
これらの方法で分析するとM. pneumoniaeは2つの系統に分かれ(図1), 遺伝子型は図1中の表のような関係になる。例えば, 実験室標準株として普及しているM129株とFH株は, ゲノム解析でT1-1とT2-Bクレードに属し, p1遺伝子型はそれぞれ1型と2型, MLVA型は4572と3662, MLST型はST1とST2である。M129株とFH株は1950~1960年代に分離された古い菌株であり, 最近分離される菌株の型は, これらとは少し異なっている。
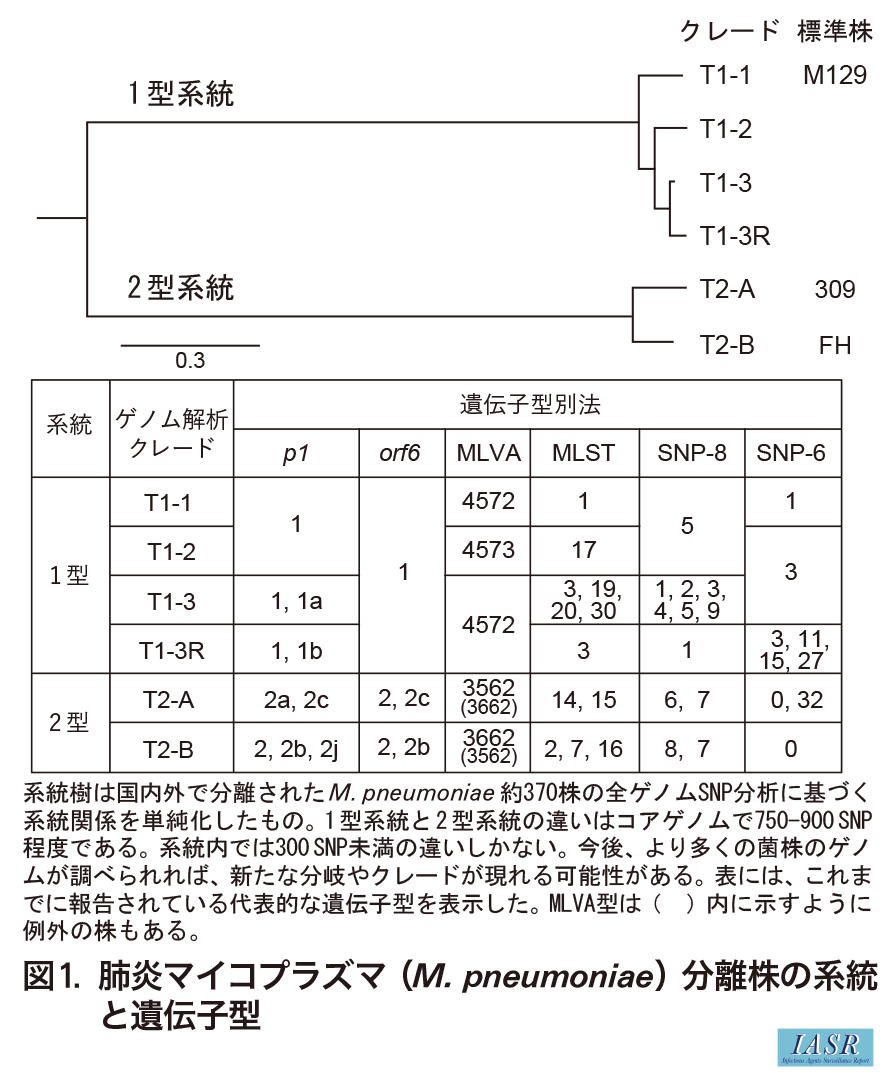{kind=link}
各遺伝子型別法にはそれぞれ特性があり, 目的によって使い分ける必要がある。菌株を細かく分類し, 感染経路の追跡を行うような目的ではMLSTやSNP分析, ゲノム解析が適している。一方, MLVA型はMLST型やSNP型よりも菌の継代によって変化しやすい。このためMLVAは単一アウトブレイクに由来する菌株の比較や鑑別には役立つが, 地理的に離れた場所や異なる時期に分離された菌株の比較には適さない。また, p1とorf6遺伝子型別法は, 分離株を細かく分類することはできないが, この菌の主要な細胞接着タンパク質, P1とP40/P90の遺伝子型であり, 宿主と病原体の相互作用を考えるうえで研究的な意義は大きい。P1とP40/P90はウイルスのスパイクタンパク質のような細胞接着分子であり, これらに生じた変異は, M. pneumoniaeが集団免疫を回避することに寄与していると考えられている6)。
M. pneumoniaeはゲノムサイズが約800kbと小さく, 現在は次世代シーケンサー(NGS)で比較的低コストで多数の株のゲノム解析が行える。ゲノム情報を取得すれば, 全ゲノムSNP分析によって菌株の詳細な系統関係がわかるとともに, MLVA型やMLST型も同時に取得できる。しかし, p1とorf6遺伝子はゲノム内に反復配列(RepMP配列)が存在するため, ショートリードNGSによるドラフト解析だけでは, p1とorf6の型は正確に決めることができない。p1とorf6の遺伝子型を正しく分析するためには, これらの遺伝子座をPCRなどで特異的に増幅した後, RFLP分析やシーケンシングによって塩基配列を確認する必要がある1,6)。
1型と2型系統の菌が検出される割合は, 調査時期によって変動がみられる。日本では, 1型と2型菌が10年程度の間隔で交互に優位になる現象が観察されている(図2)。1990年代は2型菌の分離が多かったが, 2000年代になると1型菌が優位になった。その後, 2010年代後半からは再び2型が多く分離されている。1990年代に分離されていた2型菌のp1遺伝子はFH株と同じ古い2型だが, 2010年代後半から出現している2型菌のp1遺伝子は, ほとんどが2c型か2j型で, P1とP40/P90のアミノ酸配列が少し変化している。したがって, 現在出現している2型菌は1990年代に出現していた2型菌と同じものが再出現しているのではない6)。
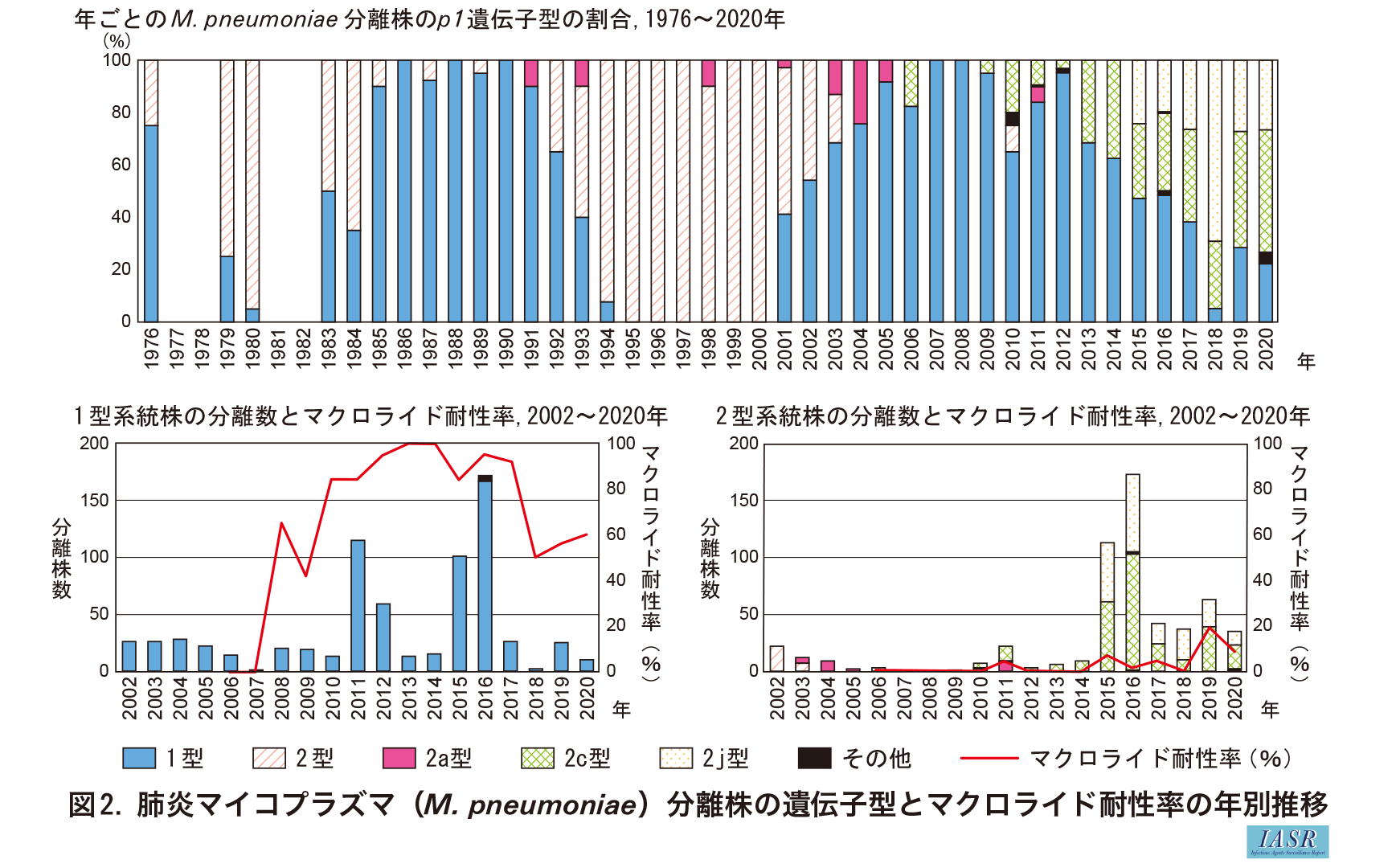{kind=link}
M. pneumoniaeの2つの系統の出現の割合は, マクロライド耐性の動向にも関連している。東アジア地域では, 2000年以降にM. pneumoniaeのマクロライド耐性化が問題となったが, 耐性菌の大部分は1型菌であった。これは1型の方が2型の菌よりも耐性化しやすいということではなく, 1型菌が2000年代に多く出現していたためであると考えられる(図2)。2000年代は分離株の耐性率が増加した時期で, 臨床でのマクロライド系抗菌薬の使用量が多かったと推測される。この時期に多く出現していた1型菌は, 2型菌よりマクロライド系抗菌薬による選択圧を受ける機会が多かったため, 2型菌より1型菌の耐性化が進んだのであろう。国内で分離されるマクロライド耐性の1型菌は, T1-3またはT1-3Rクレードに属する株が多い。一方, 国内で分離される2型菌はマクロライド耐性化があまり進んでおらず, 2010年代後半からは2型菌の出現が増えたため, 分離株全体としてマクロライド耐性率が低下している。2型菌の出現が増加した正確な要因は不明だが, 2016年からの薬剤耐性(AMR)対策アクションプラン等によって抗菌薬の使用が抑制され, 感受性菌による発症例が増えたことや, 1型菌に対して集団免疫が形成され, 流行菌が2型菌にシフトしたことなどが考えられる。一方, 現在も抗菌薬の使用量が多いとみられる中国では, 2型菌もマクロライド耐性化が進んでいることが報告されている7)。今後, 国内でも2型菌のマクロライド耐性の動向を監視するとともに, ゲノム解析によって分離株の系統関係を調べ, 伝播経路を追跡するべきであろう。
参考文献
- Cousin-Allery A, et al., Epidemiol Infect 124: 103-111, 2000
- Degrange S, et al., J Clin Microbiol 47: 914-923, 2009
- Brown RJ, et al., J Clin Microbiol 53: 3195-3203, 2015
- Touati A, et al., J Clin Microbiol 53: 3182-3194, 2015
- Zhao F, et al., iScience 24: 102447, 2021
- Kenri T, et al., Front Microbiol 14: 1202357, 2023
- Guo Z, et al., J Glob Antimicrob Resist 28: 180-184, 2022