HHV-6のヒトゲノムへの挿入 ―ciHHV-6―
(IASR Vol.41 p220-221: 2020年12月号)
HHV-6ゲノム挿入とは
HHV-6のヒトゲノムへの挿入は, 末梢血サンプルでのHHV-6感染のスクリーニングの過程で, 高コピー数のウイルスが検出されたことを契機として発見された。FISH法でヒト染色体にHHV-6ゲノムが挿入されていることが確認され, chromosomally integrated HHV-6(ciHHV-6)と呼ばれるようになった。化学療法中, 臓器移植の患者や免疫不全症のようにウイルスの再活性化が危惧される場合に同定されていたが, 現在は, 一般健常人の中に存在することがわかっており, 欧米で一般人口の1%, 日本人ではやや少なく0.21%に見つかる1)。親子, 兄弟例もあり, 生殖細胞系列を通して遺伝するので, inherited ciHHV-6(iciHHV-6)とも呼ばれている。HHV-6A/Bともに同定されるが, HHV-6Aの頻度がやや高い。
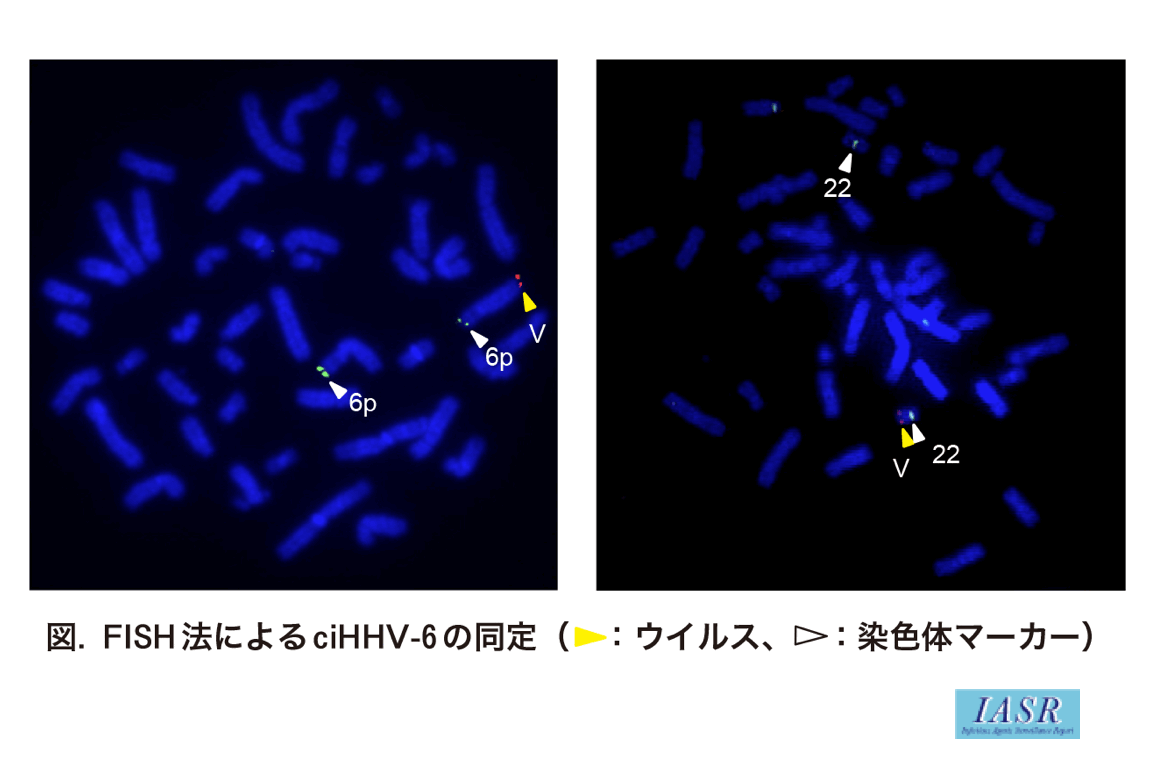
確定診断は, 末梢血サンプルでのウイルスゲノムのPCR産物をプローブとしたFISH法でなされることが多い(図)。FISH法では, 染色体に挿入されていることだけでなく, 挿入染色体の種類や挿入部位もわかる。生殖細胞系列において挿入されているので, 通常のHHV-6感染症においては検出されないような, 血液以外の爪や毛髪のような体細胞でもPCRが陽性になるので, ciHHV-6の診断に活用される。
{kind=link}
HHV-6ゲノム挿入のメカニズム
FISH法で確認すると, ほぼすべてのciHHV-6の症例で, ウイルスゲノムはテロメア領域に挿入されている。ヒトテロメア配列はTTAGGGの繰り返しで構成されているが, HHV-6ウイルスゲノムの両端にあるterminal direct repeat(DR)内にもテロメアリピート様繰り返し配列(telomere-repeat-like sequence: TRS)が2カ所ずつ存在する。ciHHV-6のゲノムDNAでは, 挿入染色体のサブテロメア配列とウイルスゲノムとにそれぞれプライマーを設計することで結合部をPCR増幅することができる。その配列解析により, ヒトゲノムとウイルスゲノムとはテロメアリピートを介して結合していることが証明された2,3)。挿入されているウイルスゲノムは必ずしも全長ではなく, また, 1コピーとは限らず複数コピーの症例もみられるが, ヒトゲノムとの接点は必ずDR内のテロメアリピートの位置である。
これらのことから, ciHHV-6におけるウイルスゲノムは, 相同組換え, もしくは相同性を利用した他のDNA修復経路によりヒトテロメア領域に挿入されていると考えられる。テロメア領域には常に宿主ゲノムDNAの断端が存在するが, ループを形成しDNA修復反応から保護されており, 相同組換えは抑制されている。一方, DNAの末端はDNA複製に不利な構造をしており, テロメアリピート長は複製回数依存性に短縮する。このようなテロメア短縮, もしくはそれ以外の何らかの理由でテロメアリピートが保護されず剥き出しになったときに, DNA末端として認識され, その時に核内に近接してウイルスゲノムが存在すると, 相同性を利用したDNA修復経路が作用し, いわば「誤修復」されることが挿入の原因であることが示唆される。
また, ヒトテロメアとウイルスゲノムが結合された後は, ウイルスゲノムが染色体末端となるわけであるが, ウイルスゲノムには都合の良いことに, 別のTRS配列も備えられており, それを染色体末端として利用して再び安定化させているようである。
ゲノム挿入は高頻度に起きているのか?
ciHHV-6は生殖細胞系列で発見されることがほとんどであるが, 腫瘍で検出された報告も散見され, 体細胞レベルでウイルス挿入は起きている。ヒト細胞株を用いて, 新規挿入を人工的に再現したデータも報告されているが, その効率は高くはない2)。挿入により細胞増殖に有利に作用するわけではないため, 通常は検出されていないようである。
興味深いことに, FISHレベルで報告されている症例報告をみると, 挿入染色体の種類は比較的限定されている。とくに22番染色体長腕(22q)末端に挿入されている症例が多い。この事実から, (1)HHV-6A/Bともに22qが多いので, HHV-6が何らかの理由で22qのテロメアに挿入されやすいのか, それとも, (2)太古の昔に起こったウイルスゲノムの挿入が, 一般集団に拡散したことによる創始者効果なのか, という疑問が生じる。そこで, 22qの末端部に近い遺伝的マーカーを利用して, 22qに挿入されているciHHV-6症例のハプロタイプ解析を行ったところ, HHV-6A/Bそれぞれで多くの症例が同じハプロタイプを示し, 共通祖先に由来することが明らかとなった4)。22q以外でも挿入染色体が比較的限定されているのも, 創始者効果を反映しているものと思われる。ゲノム挿入は実際には稀な出来事であるが, 無害なので一般集団に拡散して, いわばゲノム多型のような状態になっているようである。
ゲノム挿入は表現型に影響するのか?
ciHHV-6症例のHHV-6感染に対する免疫寛容や, 突発性発疹の臨床経過への影響などが想像できるが, そのような報告はあまりないようである。挿入部位がテロメア近傍であることもあり, 内在性レトロウイルス等と同様に, ゲノムがメチル化されて遺伝子発現が抑制されていることが予想される。ciHHV-6由来の細胞株から, ウイルスが再活性化したという報告は散見される。特殊な条件下においては, 一部のウイルス遺伝子, タンパク質が発現するのかもしれない。
一般の疾患としては, 狭心症との関連が報告されたが続報はなく, 多因子遺伝病の単なる連鎖マーカーとして作用しただけかもしれない。最近, ciHHV-6症例に習慣流産が多いというデータが得られた5)。ウイルスの再活性化が流産に関与する可能性, テロメアへの挿入が減数分裂における対合に不都合となる可能性等が考えられる。一方で, 習慣流産の罹患アリルは遺伝的適応度が低く, 淘汰されやすいことを考えると, さらに症例数を増やした検討も今後の課題である。
おわりに
ciHHV-6のゲノム挿入が塩基レベルで解明されてから10年が経つが, いつどこで何がきっかけでウイルスゲノムがテロメアに挿入されるのかは未解明のままである。種々のモデル系が利用できるようになった昨今, 今後のこの分野の研究の進展が楽しみである。
謝辞
ciHHV-6の共同研究でお世話になった藤田医科大学医学部小児科の吉川哲史教授, 河村吉紀講師, 三浦浩樹助教, 医療科学部臨床検査学科の大江瑞恵准教授に深謝致します。
参考文献
- Tanaka-Taya K, et al., J Med Virol 73(3): 465-473, 2004
- Arbuckle JH, et al., Proc Natl Acad Sci USA 107(12): 5563-5568, 2010
- Ohye T, et al., Sci Rep 4: 4559, 2014
- Kawamura Y, et al., J Gen Virol 98(7): 1823-1830, 2017
- Miura H, et al., J Infect Dis, in press