北海道における野鳥および野生哺乳動物からのH5N1亜型高病原性鳥インフルエンザウイルスの検出事例について
(IASR Vol. 43 p259-260: 2022年11月号)序 論
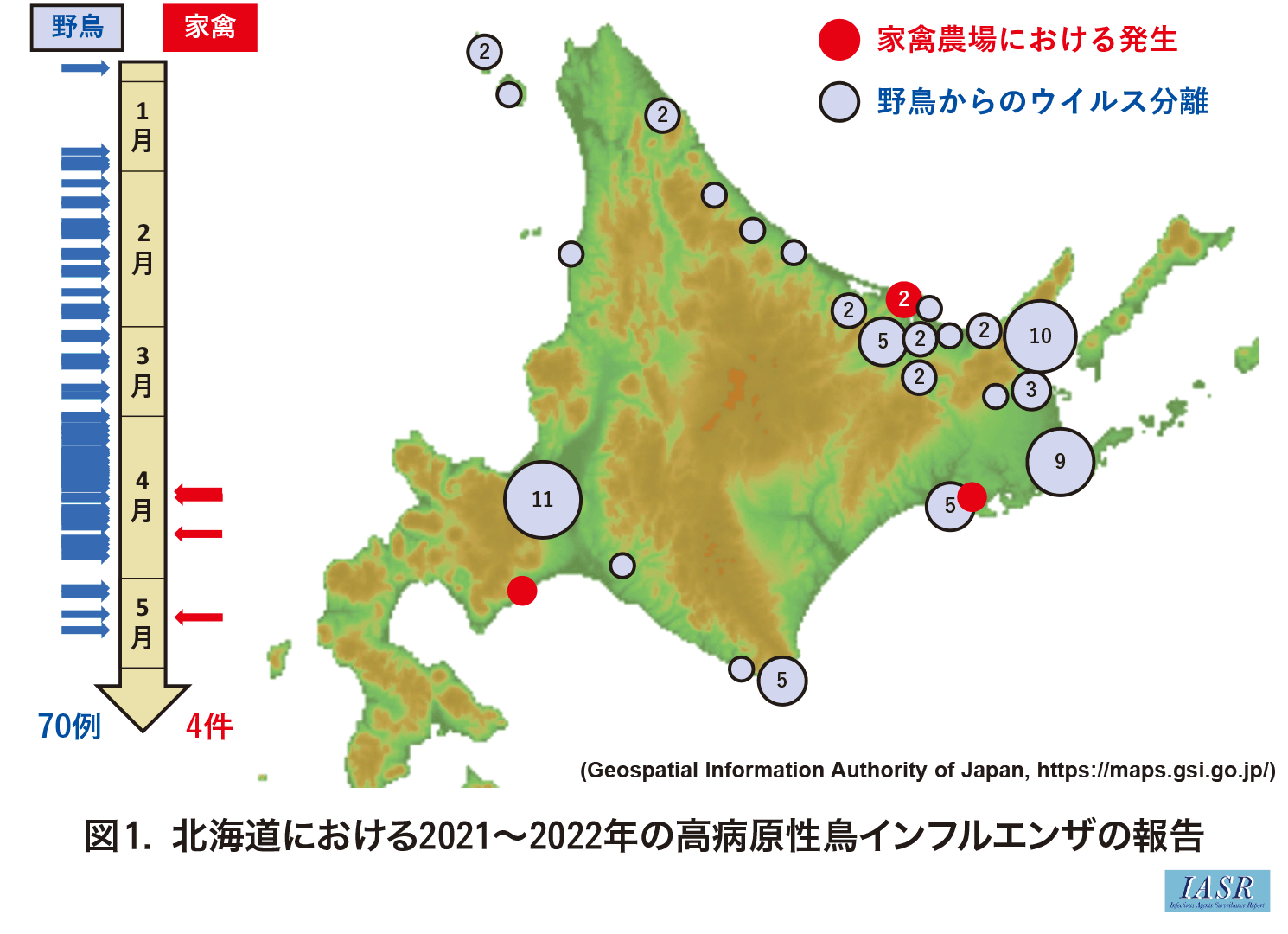
2021~2022年冬季も日本および世界で高病原性鳥インフルエンザウイルス(HPAIV)の感染事例が相次いだ。本邦では2021年11月8日~2022年5月18日までに, 全国で25件の家禽農場での発生と107例の野鳥および環境からのHPAIVの検出事例が報告され, そのうち, 北海道では4件の農場での発生と70例の野鳥からの検出が報告された(図1)1,2)。本稿では2022年1~5月までに北海道内の野鳥で確認されたHPAIV感染に関する概要を述べる。また野鳥間で循環していたと思われるHPAIVが, キタキツネやタヌキに感染した事例についても触れたい。
{kind=link}
野鳥での流行について
2022年1月2日に北海道苫前町にて発見された衰弱オジロワシからHPAIVが分離された。病原ウイルスの血清亜型はH5N1であり, これは, 2020~2021年冬季に本邦で流行したHPAIVの亜型(H5N8)とは異なるが, 2021年末から本州で検出されていたHPAIVとは同じ亜型であった3)。以降, 北海道の北部および東部で発見された死亡野鳥から同じ亜型のHPAIVが検出された3)。特筆すべきは, H5N1亜型HPAIV感染陽性として見つかった野鳥の多くはカラスと猛禽類であり, 水禽からの陽性例はほとんどなかったことである。野生水禽の渡りによるHPAIVの日本国内への侵入は広く認識されているところではあるが, 過去の発生年と比較すると野生水禽における陽性例は少なかったことから, 通常とは異なる宿主での感染例増加に関してウイルス側に特有の要因が存在している可能性がある。
3月末に, それまで北海道東部を中心に循環していたH5N1亜型HPAIVが札幌でも確認されるようになった2)。当該シーズン中, それまで札幌近郊ではH5N1亜型HPAIVの検出報告がなかったことから, 当該ウイルスの札幌圏への侵入経路については不明である。札幌市内の庭園で発見されたカラスの斃死体からH5N1亜型HPAIVの発見が初めて確認された後, 近隣の複数の庭園ならびに公園でカラスの大量死が続発した。発見斃死体から抽出し検査に供したサンプルすべてからH5N1亜型HPAIVが分離されたことから, 札幌市内の広範囲のカラスが同ウイルスの感染の影響を受けていたことが示唆される。札幌のカラスにおける継続的なH5N1亜型HPAIV感染報告とともに, 4月以降も北海道東部および南東部で発見されたカラスや猛禽類からも同ウイルスの検出例が続き, 最終的には5月14日に美幌町にて発見された死亡オジロワシからH5N1亜型HPAIVが検出された他, 我々の独自検査では5月18日に札幌市内で回収したカラスの斃死体からもウイルスが分離された3)。
分離されたH5N1亜型HPAIVの分子系統解析について
北海道内で発見された衰弱および死亡野鳥から分離されたH5N1亜型HPAIVの遺伝子配列を決定し, 分子系統解析を行ったところ, 調べたすべてのウイルスが同一の集団を形成していたことから, 北海道内への侵入時期は不明ではあるが, 同一と思われるウイルスが北海道内で循環していたことが予想される3)。これらHPAIVは, 2021~2022年冬季に欧州で流行していたウイルスの遺伝子配列と非常に近縁であることから, 由来の同じウイルスが同時期に欧州と北海道を含む極東で流行していたことが示唆された。この 1)H5N1亜型の北海道分離株, の他に, 2021~2022年冬季には2つの異なる集団のH5N1亜型HPAIVが本邦で確認されており, 2)2021年11月に鹿児島県および秋田県で発見されたHPAIVで2020~2021年冬季に西日本にて流行していたH5N8亜型ウイルスと近縁なもの, と, 3)鹿児島県や福島県で発見されたH5N1亜型HPAIVで2021年春以降に欧州で循環していたウイルスと近縁なもの, がそれぞれ異なる渡り鳥の経路によって本邦に侵入したことが予想される。北海道の家禽農場における4件の発生はすべて2022年春以降に発生しているが, そのうち2022年5月に網走の農場で検出されたH5N1亜型HPAIVは北海道内で循環していた 1)北海道分離株とは異なり, 3)鹿児島県や福島県で発見されたウイルスと近縁であった。4月以降, 本州で循環していたHPAIVが野生水禽の北方への渡りを介し北海道に持ち込まれたことが予想される。
野生哺乳動物におけるHPAIV感染事例について
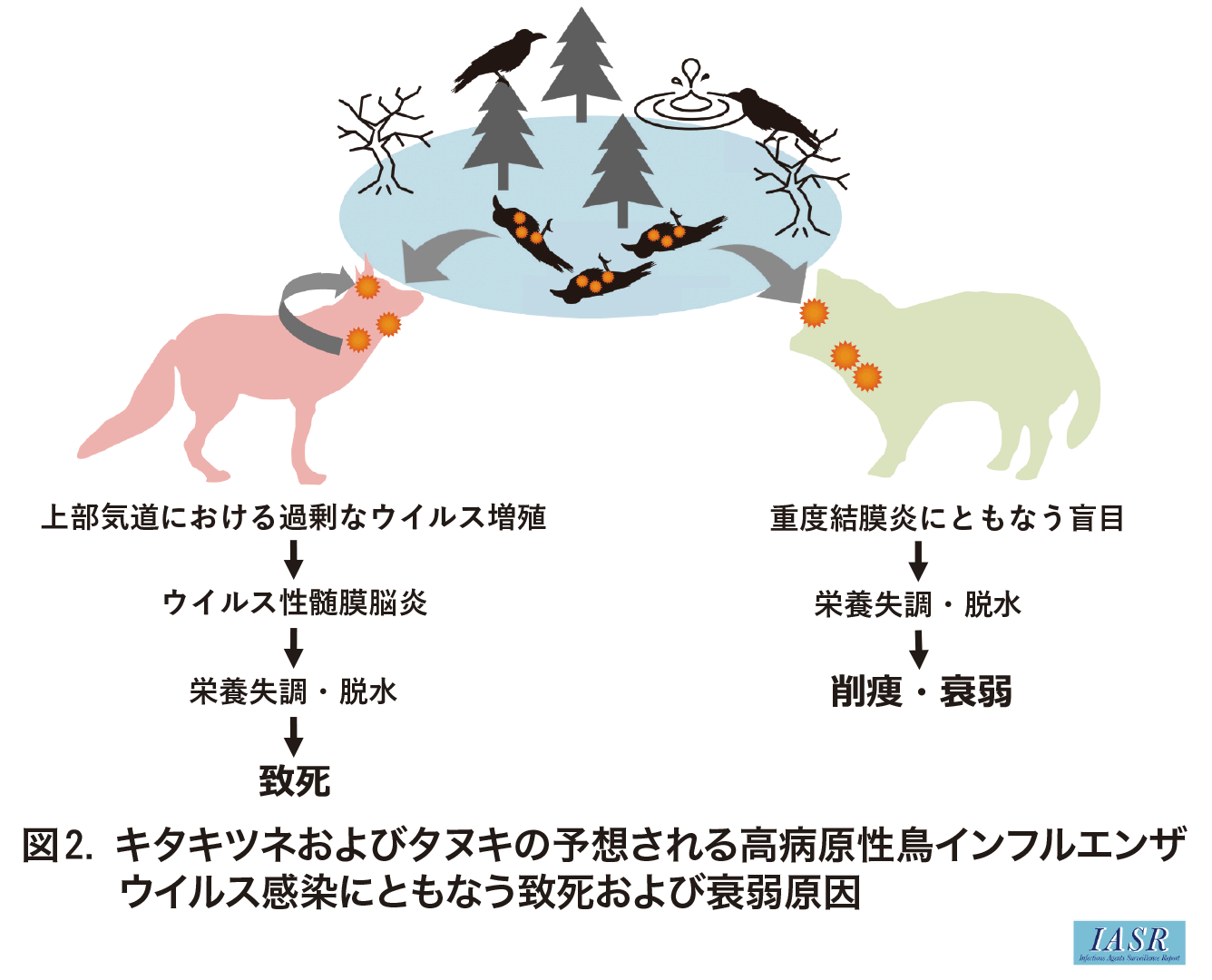
札幌市内でカラスからHPAIVが初めて検出された際に, 同庭園にて斃死キタキツネと衰弱タヌキが確認された。適切な処理後に臓器乳剤を作製し10日齢発育鶏卵に接種したところ, キタキツネおよびタヌキともに脳乳剤材料からHPAIVが分離された4)。分離ウイルスの遺伝子配列は, 上記1)北海道分離株とほぼ同じであり, polymerase basic protein 2(PB2)の627番目や701番目, およびhemagglutinin protein(HA)の226番目や228番目などの哺乳動物への馴化に寄与するアミノ酸置換は認められなかった5-7)。
各動物におけるHPAIV感染の詳細な病態を調べるために病理組織学的検索を行った。キタキツネの気管や気管支ではHPAIV抗原が検出されたことから, キタキツネはH5N1亜型HPAIVに呼吸器感染したことが強く疑われる。さらにウイルスが分離された脳における病理組織学的検索では, 髄膜を中心に好中球の浸潤をともなう軽度の炎症像が認められたことから, キタキツネは上部気道にてHPAIV感染を引き起こし, その後, ウイルス性髄膜脳炎を引き起こしたと思われる(図2)。一方, タヌキでも気管においてH5N1亜型HPAIV抗原が確認され, 好中球の他にリンパ球や形質細胞の浸潤をともなう慢性反応性の炎症像を認めた。さらにタヌキでは線維化や細菌の二次感染をともなうウイルス性結膜炎を認めたことから, 盲目による削痩で衰弱していたことが示唆された。キタキツネによるカラスの捕食は広く知られていることから, 感染カラスの捕食によるキタキツネのHPAIV感染が疑われる一方, タヌキは盲目下で感染カラスに偶発的に接触したことが予想される。
{kind=link}
終わりに
北海道における2021~2022年冬季のH5N1亜型HPAIV感染は通常と異なる宿主動物から多く報告されていたことから, 近年循環しているH5亜型HPAIVの遺伝子および病原性をより詳細に解析する必要がある。またこのような, HPAIVの病原性や宿主域の変化により, 渡り鳥によって運ばれるHPAIVの疫学が大きく変化している可能性もある。引き続き野外で検出したHPAIVの遺伝子および病原性に関する情報公開を進める必要がある。
参考文献
- 農林水産省, 鳥インフルエンザに関する情報
https://www.maff.go.jp/j/syouan/douei/tori/ - 環境省, 高病原性鳥インフルエンザに関する情報
https://www.env.go.jp/nature/dobutsu/bird_flu/ - Isoda N, et al., Viruses 14: 2168, 2022
- Hiono T, et al., Virological, pathological, and glycovirological investigations of an Ezo red fox and a tanuki naturally infected with H5N1 high pathogenicity avian influenza viruses in Hokkaido, Japan. Under article submission
- Hatta M, et al., Science 293: 1840-1842, 2001
- Li Z, et al., J Virol 79: 12058-12064, 2005
- Rogers GN & Paulson JC, Virology 127: 361-373, 1983