日本紅斑熱リケッチア媒介マダニの国内分布状況
(IASR Vol. 41 p138-139: 2020年8月号)
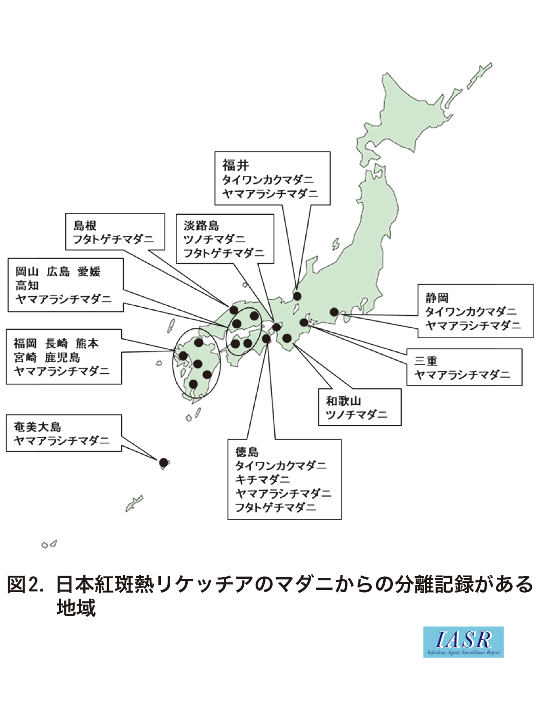
国内の日本紅斑熱リケッチアRickettsia japonicaの媒介種として, これまでに3属8種がリストアップされており1), このうちR. japonicaの分離は, タイワンカクマダニ, ツノチマダニ, キチマダニ, ヤマアラシチマダニおよびフタトゲチマダニの5種から, 遺伝子検出のみはタカサゴチマダニ, オオトゲチマダニおよびヤマトマダニの3種からとされている。これらマダニ種の地理的分布を図1に, リケッチアが分離された地域とマダニの種類を図2に示した。これら8種は大まかに2つの分布パターンに分けられる。タイワンカクマダニ, ヤマアラシチマダニ, タカサゴチマダニ, そしておそらくツノチマダニの4種は日本紅斑熱(以下, 紅斑熱)発生地域と重なり合うように千葉県以西へ偏る傾向があるが, 他の4種は全国分布である。
{kind=link}
{kind=link}
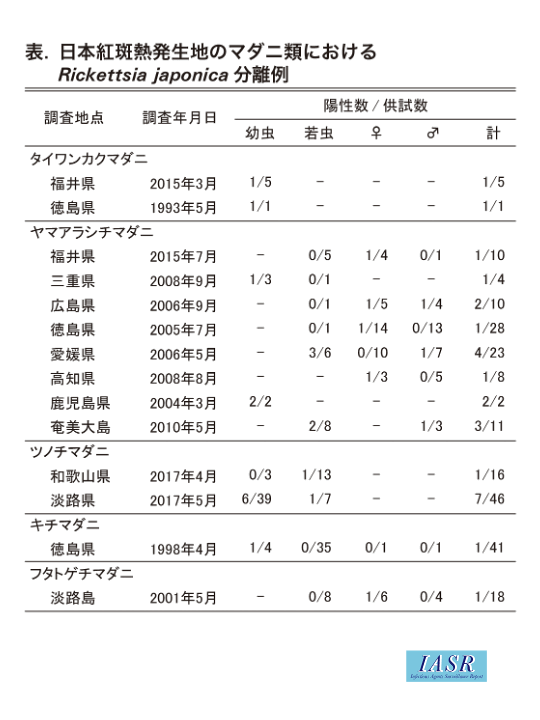
これらのマダニ種と紅斑熱発生との具体的関係については不明な部分が多い。マダニ類の病原体保有の程度は全国一律ではないようであり, 媒介種と目されているマダニの生息自体が紅斑熱感染のリスクに直結しているとも思えない。なぜならば, R. japonicaは, 厳密には特定のマダニ種の共生体ではないと推測されるからである。共生体の条件はマダニの種類に特化したほぼ100%の保有率であり, これら8種にはそのような関係を持つ種類はない。ただし, 紅斑熱発生地には, R. japonicaに親和性が高く, これと共生的関係を維持しているマダニ種の家系の散在が推測される。図1は, 紅斑熱発生地におけるマダニ類からのR. japonica分離成績の一部である。ここでの供試数は1回の調査において採集し得たほぼ全数に相当する。供試数がまだ少数にとどまっている地点が多いため, 地点間における分離率の比較は難しいが, 図2と合わせると各地のR. japonica保有種の概要はうかがえるかと思う。
タイワンカクマダニ, ヤマアラシチマダニおよびツノチマダニにおいては, 経卵感染の傍証となる未吸血幼虫からのリケッチア検出例も複数回知られることから, R. japonicaとの親和性の高さがうかがわれ, 媒介種としての可能性は高いと考えられる。これら3種の刺咬にかかわるヒト嗜好性は他種に比べて低いことが指摘されているが, 人体刺咬症例はいずれの種類においても記録されている(表)。
{kind=link}
紅斑熱発生地のマダニ類における長年の分離検査にもかかわらず, キチマダニとフタトゲチマダニの未吸血個体からのR. japonicaの分離はそれぞれ1例のみ, タカサゴチマダニからの分離は皆無, オオトゲチマダニとヤマトマダニからの分離株はすべてR. japonica以外のリケッチアである(表)。
フタトゲチマダニは単為系と両性系が存在し, 紅斑熱発生地域は両性系もしくは単為・両性系の混在地域に偏る傾向がある。紅斑熱患者から回収されたフタトゲチマダニからのR. japonica分離が島根県2)と徳島県から各1例知られるが, このときの虫体がR. japonicaを保有していて媒介したとは必ずしも断定できず, もともと未保有であったマダニが既感染患者から偶発的にリケッチアを取り込んだ状態で回収された可能性は否定できない。淡路島の1分離例のみが植生上から採集された未吸血虫体に由来する。フタトゲチマダニは, 最も多数の虫体がリケッチア検査に供されてきた種類であるが, これまでに遺伝子検出・分離からはともにR. japonica近似種のRickettsia sp. LONが大多数を占める。例外的に, 島根県の紅斑熱多発地において比較的高いR. japonica遺伝子の検出率が報告されたことがある2)。
R. japonicaは比較的分離が容易とされることから, 遺伝子が検出されたマダニ種の生息地域においては, 同種マダニからのリケッチア分離が期待されてきたが, 未だにタカサゴチマダニ, オオトゲチマダニおよびヤマトマダニからの分離例は皆無で, またキチマダニとフタトゲチマダニにおいても, これまでの供試数の多さからはほぼ分離率ゼロに等しい。遺伝子検出陽性例だけで分離ができていないマダニ種が存在する事実は, 媒介種のさらなる解明にとっての今後の検討課題である。
参考文献
- 安藤秀二ら, 衛生動物 64: 5-7, 2013
- Tabara K, et al., J Vet Med Sci 73: 507-510, 2011