新型コロナウイルスSARS-CoV-2 Spikeタンパク質 E484K変異を有するB.1.1.316系統の国内流入(2021年2月2日現在)
新型コロナウイルス変異株が有するSpikeタンパク質E484K変異
自治体の積極的疫学調査を支援すべく、SARS-CoV-2(一本鎖プラス鎖RNAウイルス、全長29.9 kb)のゲノム配列を確定してGISAID*1へ配列登録し、感染クラスターに特有な遺伝子情報およびクラスター間の共通性を解析中である。これまでに4回にわたって中間報告として国内伝播の状況を概説してきた(2020年4月27日1)、2020年8月6日2)、2020年12月11日3)、2021年1月14日4))。2020年3~4月に欧州系統(Pangolin*2系統B.1.1.114)の流入が認められたものの、第2、3波の主流はこの欧州系統から派生したB.1.1.284とB.1.1.214による国内の感染拡大であることが判明した(2021年1月14日報告4))。さらに、英国で発生した新規変異株VOC-202012/01 の流入が懸念されており、空港検疫検査のみならず、市中感染の疑いを示す事例が公表されている。
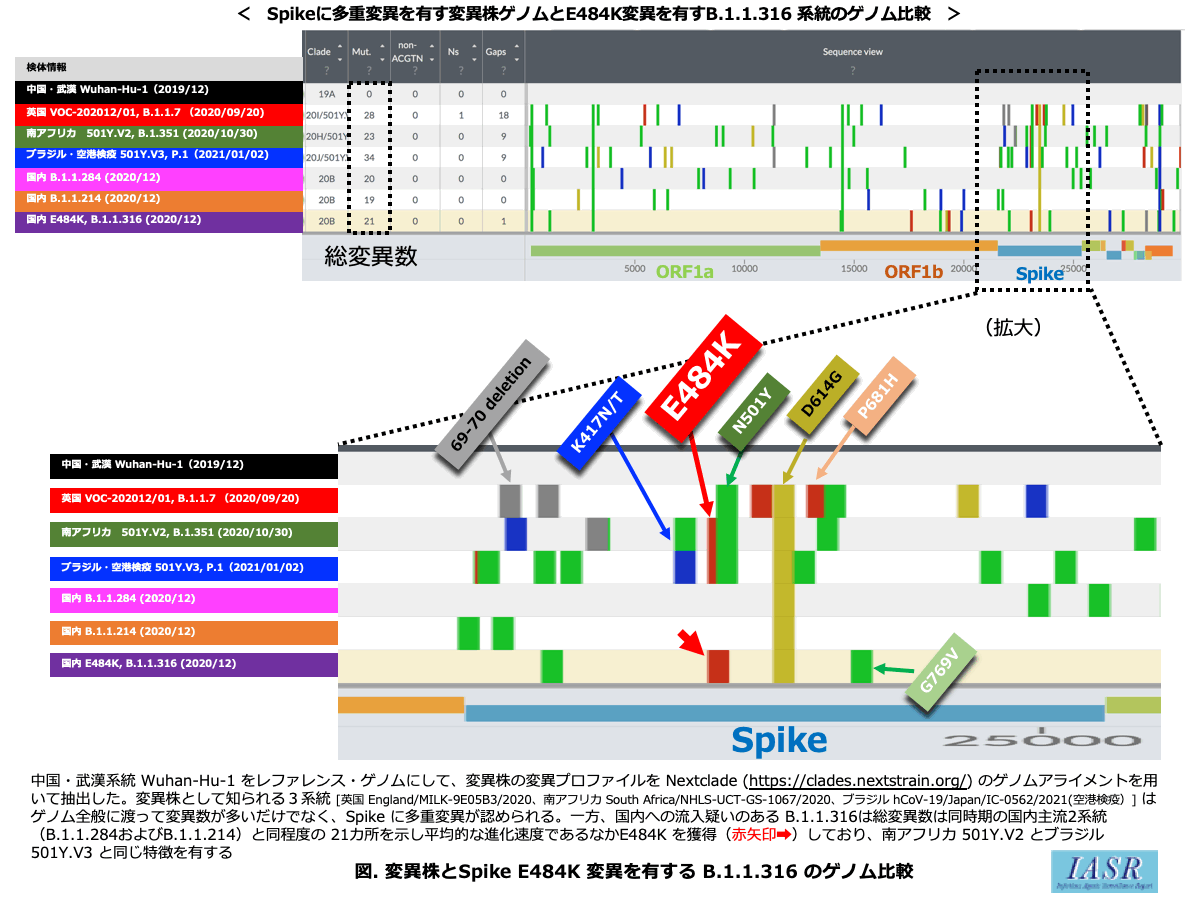
Spikeタンパク質の多重変異を特徴とする変異株は発生国を基に3系統〔英国VOC-202012/01(B.1.1.7)、南アフリカ501Y.V2(B.1.351)、ブラジル501Y.V3(P.1)〕が報告されている。変異株は感染伝播力の上昇およびワクチン効果を減弱させる免疫逃避の可能性が指摘され、国内への流入と感染伝播を注視している。変異株の特徴の詳細は国立感染症研究所ホームページのリスクアセスメント報告「感染・伝播性の増加や抗原性の変化が懸念される 新型コロナウイルス(SARS-CoV-2)の新規変異株について(第5報)」をご参照いただきたい5)。
Spikeタンパク質はコロナウイルスの表面を覆い、ヒト受容体(新型コロナの場合はACE2タンパク質)に結合する。その結合領域(receptor-binding domain: RBD、328–533aa)に親和性を示す中和抗体が感染防御に最も有効であることが知られている6)。変異株はRBDの2-3カ所に特徴的な変異を有し(N501Y、E484K、K417T/N)、特にアミノ酸484番目のグルタミン酸(E)はRBDのACE2結合に重要であり、かつ中和抗体の中心エピトープに配置されるアミノ酸残基である。グルタミン酸(E)は陰性荷電を示す酸性極性側鎖アミノ酸である一方、リジン(K)は陽性荷電を示す塩基性アミノ酸に分類される。つまり、E484K変異は磁石のNS極を逆転させるような極性を反転させる際立った変異であり着目に値する。このE484K単独でワクチンは無効化されないが、少なからず効果が減弱する可能性が指摘されている7-9)。
Spikeタンパク質E484K変異を有する系統(Pangolin B.1.1.316)の国内拡散
COVID-19ゲノムサーベイランス*3の全国調査において、検体採取日が2020年12月上旬以降から南アフリカやブラジルで報告されている変異株(N501Y.V2とN501Y.V3)と同一のSpikeタンパク質にE484K変異を有するB.1.1.316系統(空港検疫2件、関東全域91件)を検出した(2021/02/02現在)。B.1.1.316系統は、感染性・伝播性の高まる可能性のある変異であるN501Y変異は有しておらず、N501Y変異を有する変異株とは異なる表現型であることが推測される。ゲノムネットワーク図の評価では、このB.1.1.316は少なくとも現在の国内の主流2系統の系譜ではなく、2020年3~4月・欧州系統(B.1.1.114)から13塩基変異(およそ7カ月間の時間差)を有していた。国内検体でこの13塩基変異の空白リンクを埋める検体は特定されておらず、国内で変異を獲得した系譜ではないと判断された。また、GISAID全登録ゲノム情報を検索しても、この流入B.1.1.316株の起源を示す発生国を特定できていない(つまり、英国等地域を断定できない)。一方、国内の主流2系統(B.1.1.284およびB.1.1.214)からE484K変異を獲得した株は現在のところ検出されていない。
{kind=link}
おわりに
国内においてもワクチン接種が開始される。感染伝播を抑制する重要なツールであるが、ワクチン導入後のウイルスの適応変異に関しても、十分に注意する必要がある。これまでの国内の感染伝播を抑える努力とともに、世界的に懸念されている株の国内流入、あるいは国内での変異株の出現の早期探知も同時に重要となる。これらに対応するためにも、体系的で継続的なゲノムサーベイランスの確立が重要である。
*1 Global Initiative on Sharing All Influenza Data (GISAID)10)。
GISAIDは、鳥インフルエンザが猛威をふるった2006年8月に医療分野の研究者たちによって設立されたインフルエンザウイルスの情報データベースである。SARS-CoV-2ゲノム情報もGISAIDが主体的に運用し、登録・収集されている。毎年一定数の検体から病原体遺伝子情報を取得して歴年の発生動向を調査し、伝播状況やワクチン株選定等に利活用される。~1,000株/年ほどの日本のインフルエンザウイルス(A/H1およびA/H3)のワクチン抗原(ヘマグルチニンHA)および抗インフルエンザウイルス薬阻害ターゲット(ノイラミニダーゼNA)の遺伝的特徴が登録されている。
*2 COVID-19 Lineage Assigner Phylogenetic Assignment of Named Global Outbreak LINeages (Pangolin)
新型コロナウイルスのゲノム情報を元にした分子系統解析ID (https://cov-lineages.org/lineages.html)
*3 COVID-19 ゲノムサーベイランス
本調査は地域バイアスを伴った評価であることがぬぐえない。このため、できうる限り直近の陽性検体においてさらなる追加調査が必須であると考える。
謝辞:検体採取等調査にご協力いただきました医療機関、保健所および行政機関の関係者に深謝致します。
本研究は日本医療研究開発機構AMED(研究課題番号:JP19fk0108104,JP20fk0108103)と厚生労働行政推進調査事業費・「新型コロナウイルス感染症等の感染症サーベイランス体制の抜本的拡充に向けた人材育成と感染症疫学的手法の開発研究」(班長・鈴木 基、分担・黒田 誠)の研究支援を受け実施した。
本稿に関連し、開示すべき利益相反状態にある企業等はありません。
参考文献
- 新型コロナウイルスSARS-CoV-2のゲノム分子疫学調査
https://www.niid.go.jp/niid/ja/basic-science/467-genome/9586-genome-2020-1.html(2020) - 新型コロナウイルスSARS-CoV-2のゲノム分子疫学調査2 (2020/7/16現在)
https://www.niid.go.jp/niid/ja/basic-science/467-genome/9787-genome-2020-2.html(2020) - 新型コロナウイルスSARS-CoV-2のゲノム分子疫学調査(2020年10月26日現在)
https://www.niid.go.jp/niid/ja/diseases/ka/corona-virus/2019-ncov/2488-idsc/iasr-news/10022-491p01.html(2021) - 新型コロナウイルスSARS-CoV-2ゲノム情報による分子疫学調査(2021年1月14日現在)
https://www.niid.go.jp/niid/ja/diseases/ka/corona-virus/2019-ncov/2488-idsc/iasr-news/10152-493p01.html(2021) - 感染・伝播性の増加や抗原性の変化が懸念される 新型コロナウイルス(SARS-CoV-2)の新規変異株について (第5報)
https://www.niid.go.jp/niid/ja/diseases/ka/corona-virus/2019-ncov/10144-covid19-34.html(2021) - Barnes, C.O., et al., SARS-CoV-2 neutralizing antibody structures inform therapeutic strategies, Nature 588, 682-687, doi:10.1038/s41586-020-2852-1(2020)
- Gard Nelson, O.B., Patricia Spilman, Kayvan Niazi, Shahrooz Rabizadeh, Patrick Soon-Shiong, Molecular dynamic simulation reveals E484K mutation enhances spike RBD-ACE2 affinity and the combination of E484K, K417N and N501Y mutations (501Y.V2 variant) induces conformational change greater than N501Y mutant alone, potentially resulting in an escape mutant, bioRxiv, doi:https://doi.org/10.1101/2021.01.13.426558(2021)
- Sonia Jangra, C.Y., Raveen Rathnasinghe, Daniel Stadlbauer, PVI study group, Florian Krammer, Viviana Simon, Luis Martinez-Sobrido, Adolfo García-Sastre, Michael Schotsaert, The E484K mutation in the SARS-CoV-2 spike protein reduces but does not abolish neutralizing activity of human convalescent and post-vaccination sera, medRxiv, doi:
https://doi.org/10.1101/2021.01.26.21250543(2021) - Wang, Z. et al., mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants, Nature, doi:10.1038/s41586-021-03324-6(2021)
- https://www.gisaid.org/epiflu-applications/next-hcov-19-app/